一种铁腐蚀辅助的供氢系统:低氢分压下产甲烷菌与产乙酸菌的培养方法
来源:武汉市灰藻生物科技有限公司 浏览量:29 发布时间:2026-06-29 22:29:31
摘要
氢气(H₂)是厌氧环境中的重要发酵中间产物。尽管自然环境中的氢分压极低,但氢利用型微生物的培养与分离通常需要在极高的氢分压下进行,这可能会阻碍我们对适应低氢环境微生物的发现与认知。
在此,我们构建了一种被称为"铁腐蚀辅助供氢(iCH)系统"的培养体系。该系统通过连通两个培养瓶(一个用于铁腐蚀反应,另一个用于微生物培养)的气相,实现了在低氢分压下的微生物培养。 我们以水稻田土壤为微生物来源,对产甲烷菌和产乙酸菌进行了富集培养。
在传统的高氢分压富集培养中,仅富集到了Methanobacterium(甲烷杆菌属);相比之下, 适应低氢环境的产甲烷菌Methanocella(甲烷胞菌属)和Methanoculleus(甲烷微菌属)在iCH培养中被特异性地富集。
此外,我们还观察到iCH系统对产乙酸菌具有选择性富集作用(如Acetobacterium醋酸杆菌属和Sporomusa孢八叠球菌属),而在高氢培养中则以Clostridium(梭菌属)占优势。 这些结果表明,iCH系统有助于在低氢分压下培养厌氧微生物,从而能够实现对适应低氢环境微生物的选择性培养。
引言
分子氢(H₂)是厌氧环境中重要的中间代谢物和能量载体。由于H₂在自然厌氧环境中周转极快,其分压通常极低,仅为几帕至几十帕(Pa)。然而,在常规研究中,氢利用型微生物的培养与分离通常在高氢分压(100 kPa或更高)下进行。 在这种实验室条件下,很难得出关于氢利用型微生物在自然环境中生态生理学的确切结论,也无法分离出适应低氢环境的微生物。
事实上,通过宏基因组学等分子环境分析手段已经推断, 在那些预估氢浓度极低(即有机质可利用性低的环境,包括地下环境、泥炭土和深海沉积物)的厌氧环境中,存在着尚未被培养的氢利用型产甲烷菌和产乙酸菌。
由于低分压供应的H₂会被迅速消耗,常规的批次培养系统无法获得充足的微生物生长。迄今为止,已有几个研究团队开发了能在低氢分压下连续供氢的方法,以阐明低氢环境中氢营养型产甲烷菌的生态生理学。
Morgan等人利用连续通入含H₂混合气体的连续培养系统,实现了氢营养型产甲烷菌的低氢培养。利用该系统,作者发现产甲烷途径中某些代谢酶的表达受H₂浓度的调控。 类似的方法在后续关于氢营养型产甲烷菌低氢响应的研究中被频繁采用。
Sakai等人开发了"共培养法",即将产甲烷菌与异养产氢细菌共培养,以实现低浓度H₂的持续供应。 该方法实现了对预期适应低氢分压的未培养氢营养型产甲烷菌的选择性富集,并最终成功分离出在系统发育上具有新颖性的产甲烷菌,如甲烷胞菌属(Methanocella spp.)和甲烷线菌属(Methanolinea spp.)。 共培养法也被用于分析产甲烷菌对低氢分压的生理响应。
尽管这些方法成功实现了低氢分压下的实验室培养,但仍有一些问题亟待解决。连续通气过程由于需要相对复杂的系统(包括大型或小型反应器及供气装置), 无法与大量培养平行进行。虽然共培养法只需简单的系统,适合用于富集培养,但由于其依赖于与发酵细菌的共存,因此不能直接用于氢利用型微生物的分离。 此外,也不能排除除H₂以外的其他代谢物(如乙酸等有机酸)对氢利用型微生物生长的影响。
在本研究中,我们旨在开发一种能够在低氢分压下对微生物进行选择性培养的简便方法。我们关注的核心反应是金属铁在无氧溶液中的腐蚀:Fe⁰ + 2H⁺ ↔ Fe²⁺ + H₂。 利用铁腐蚀产生的H₂来培养微生物的概念此前已有报道。相关作者证明,氢营养型产甲烷菌可以利用金属铁产生的H₂作为唯一能源进行培养。
然而,该方法此前尚未被应用于培养适应低氢环境的微生物。 考虑到在无氧且接近中性的溶液中,由于Fe⁰氧化(E⁰ʹ ≈ −0.47 V)与质子还原生成H₂(E⁰ʹ ≈ −0.41 V)的标准氧化还原电位差值较小,铁的腐蚀进行得非常缓慢,因此可以预期, 通过铁腐蚀供氢非常适合在低氢分压下培养氢营养型微生物。在此,我们报道了一种基于铁腐蚀反应的培养系统,该系统已成功用于氢营养型产甲烷菌和产乙酸菌的选择性富集,这些微生物具有适应极低氢含量环境的潜力。
一、材料与方法
1、铁腐蚀持续供氢系统构建
由于微生物在低氢分压下的生长通常非常缓慢,培养系统需要能够长时间持续供应氢气。因此,我们首先验证了铁腐蚀反应能否实现氢气的长期连续供给。此外,为了开发一种能够调节氢气供应速率的培养系统, 我们在无氧缓冲液中分别添加了不同粒径和用量的零价铁(Fe⁰)(包括直径 1–2 mm 的 Fe⁰ 颗粒,以及直径 < 45 µm 的 Fe⁰ 粉末),并测定了相应的氢气产生速率(图 1)。 构建包含两个培养瓶的iCH系统:左侧培养瓶用于微生物培养,右侧培养瓶用于铁腐蚀(即产氢)反应。两个培养瓶的气相通过不锈钢管相连,使腐蚀瓶中产生的氢气得以扩散。(图 2)
2、铁腐蚀持续供氢的验证
我们在添加零价铁(Fe⁰)颗粒的培养瓶中观察到氢气连续产生超过 6 个月(图 1A)。产氢速率几乎与 Fe⁰ 颗粒的添加量(每瓶 0.03 至 3 g)成正比,范围在每瓶每天 0.12 至 4.4 μmol 之间(图 1C)。 使用 Fe⁰ 粉末时,在添加量为每瓶 3 g、1 g 和 0.3 g 的培养瓶中,产氢分别在约 2 周、1 个月和 3 个月后停止(图 1B)。氢气的最高积累量约为每瓶 1000 μmol,相当于约 50 kPa。 我们认为产氢停止是出于热力学原因(氢分压的增加和质子浓度的降低)。产氢速率同样与 Fe⁰ 粉末的添加量成正比,范围为每瓶每天 2.0 至 93 μmol(图 1C)。由于球形材料的比表面积与其直径成反比, 假设所用颗粒为球形,Fe⁰ 粉末(< 45 µm)的表面积是 Fe⁰ 颗粒(1–2 mm)的 20 倍。Fe⁰ 粉末的产氢量是 Fe⁰ 颗粒的 16 至 22 倍(图 1C), 这表明表面积是决定产氢速率的主要因素。这些结果表明,利用铁腐蚀反应可以实现氢气的长期供应,并且可以通过改变 Fe⁰ 颗粒的粒径和用量(即总表面积)来调节氢气的供应速率。
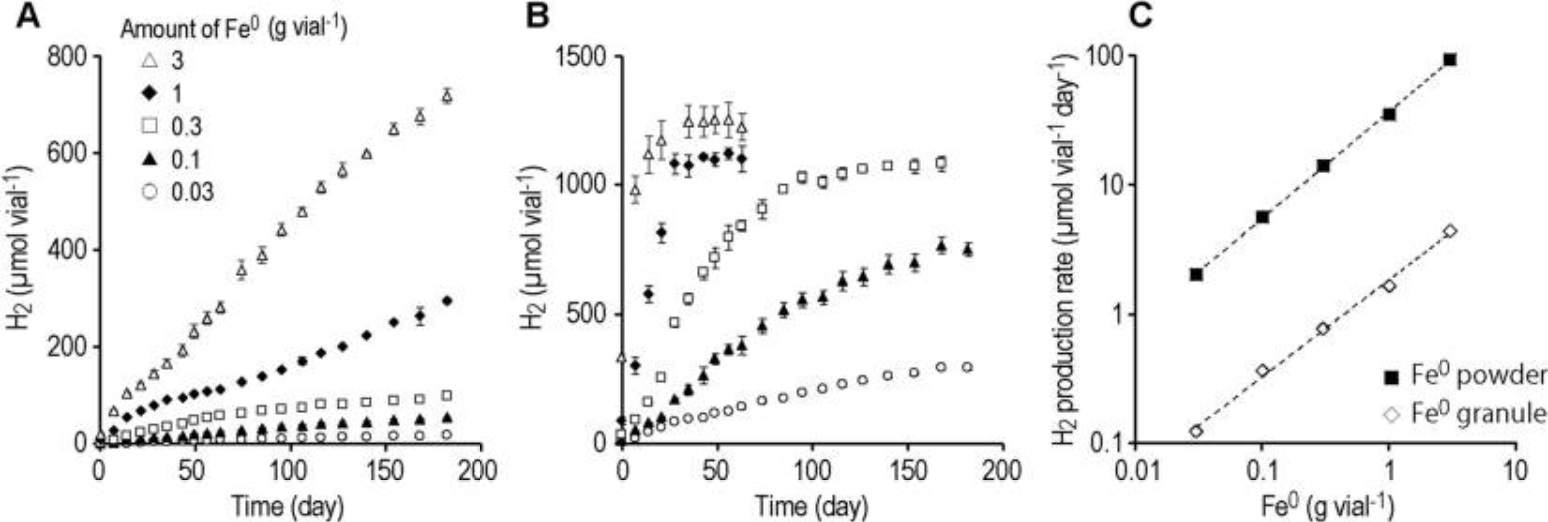
图1:零价铁(Fe⁰)颗粒(1–2 mm)(A)和 Fe⁰ 粉末(< 45 µm)(B)通过腐蚀反应连续产氢。(C)根据 A 和 B 中的数据计算得出的产氢速率

图2:铁腐蚀辅助供氢(iCH)系统示意图。左侧培养瓶用于微生物培养,右侧培养瓶用于铁腐蚀(即产氢)反应。两个培养瓶的气相通过不锈钢管相连,以使腐蚀瓶中产生的氢气得以扩散。
3、微生物培养实验
3.1、纯培养实验
以甲酸甲烷杆菌(Methanobacterium formicicum)作为模式菌株,在iCH系统中接种,并以每瓶1 g的零价铁(Fe⁰)粉末作为氢源,监测气相中CH₄和H₂的含量。
3.2、富集培养实验
以稻田土壤为微生物来源,设置三种培养条件:
01、高氢富集:气相中H₂分压为160 kPa
02、Fe⁰富集:微生物与每瓶1 g的Fe⁰粉末在同一瓶中培养
03、iCH富集:采用铁腐蚀辅助供氢系统
4、分析方法
4.1、代谢产物分析
所有代谢物的量均使用各自的半反应方程式,以电子当量(单位为mmol e⁻/培养瓶)表示:
2H⁺ + 2e⁻ ↔ H₂
HCO₃⁻ + 9H⁺ + 8e⁻ ↔ CH₄ + 3H₂O
2HCO₃⁻ + 9H⁺ + 8e⁻ ↔ CH₃COO⁻ + 4H₂O
4.2、微生物群落分析
通过对16S rRNA基因扩增子进行高通量测序分析,评估富集培养物和接种土壤的微生物群落结构。操作分类单元(OTUs)以97%的序列相似度为阈值进行划分。
二、结果与讨论
1、iCH系统中氢营养型产甲烷菌的纯培养
我们以氢营养型产甲烷菌——甲酸甲烷杆菌(Methanobacterium formicicum)作为模式菌株,评估了 iCH 系统在低氢分压下培养微生物的能力。该菌株已知在高、低氢分压条件下均能生长。 我们在 iCH 系统中接种了 M. formicicum,并以每瓶 1 g 的零价铁(Fe⁰)粉末作为氢源,同时监测了气相中 CH₄ 和 H₂ 的含量(图 3)。我们观察到, 在 iCH 系统中,M. formicicum 在近 1 个月的时间内持续产生 CH₄。尽管在培养初期出现了 H₂ 的积累(第 3 天时约为 190 Pa,相当于每瓶 7.4 µmol),但随后 H₂ 分压下降, 并在第 14 天后维持在极低水平(30 至 50 Pa)。所观察到的 H₂ 分压与天然厌氧环境以及产甲烷菌与发酵细菌的实验室共培养体系中观察到的分压相当。这些结果表明,iCH 系统能够在低氢分压下实现氢利用型微生物的长期培养。
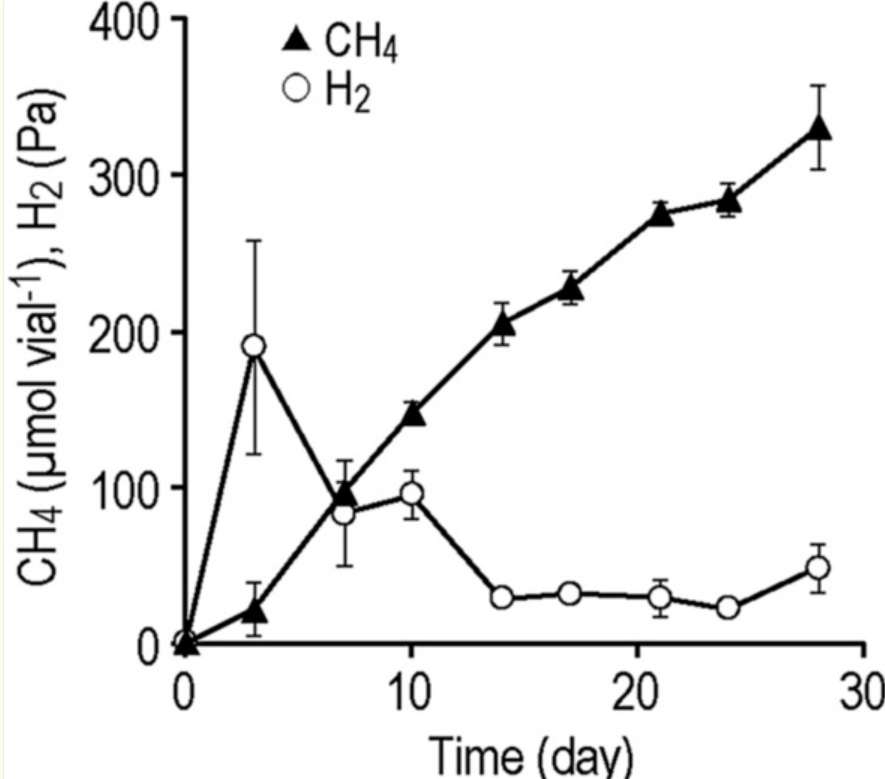
图3:在添加 1 g 零价铁(Fe⁰)粉末的铁腐蚀辅助供氢(iCH)系统中培养甲酸甲烷杆菌(Methanobacterium formicicum)期间的甲烷(CH₄)产量与氢气(H₂)分压。数据以三次独立培养的平均值表示,误差棒代表标准偏差。
2、利用 iCH 系统进行氢营养型产甲烷菌的富集培养
为了证明 iCH 系统能够选择性地培养适应低氢分压的微生物,我们进行了产甲烷菌的富集培养实验。我们选用稻田土壤作为微生物来源,因为已知稻田土壤是一种低氢环境, 并且也是已知能适应低氢环境的甲烷胞菌属(Methanocella spp.)的分离来源。除了 iCH 系统(以每瓶 1 g 的零价铁(Fe⁰)粉末作为氢源)外, 我们还设置了高氢分压下的常规培养(气相中 H₂ 分压为 160 kPa,以下简称为“高氢富集”)以及铁腐蚀条件下的培养(微生物与每瓶 1 g 的 Fe⁰ 粉末在同一瓶中培养,以下简称为“Fe⁰ 富集”)作为对照实验。
我们在添加了利福平以抑制细菌生长的无机培养基中,对氢营养型产甲烷菌进行富集。经过三次连续传代培养后,我们在培养期间分析了代谢产物(CH₄ 和 H₂)(图 4A–C)。在高氢富集组中, 我们观察到的 CH₄ 产量几乎等于根据 H₂ 消耗量计算出的理论值(图 4A)。在 Fe⁰ 富集组中,在培养初期(第 0–14 天),我们观察到的 CH₄ 产量与根据铁腐蚀产氢量计算出的理论值(图 4B 中的虚线)相当; 然而,在培养后期(第 14 天之后),CH₄ 产量趋于平稳。在培养期间,Fe⁰ 富集组培养液的 pH 值从 7.0 升高至 8.1 左右,而其他富集组的培养液 pH 值保持在 7.0 左右。 pH 值的升高,以及可能伴随的亚铁离子浓度的增加,可能抑制了 Fe⁰ 富集组中产甲烷菌的生长。在 iCH 富集组中,CH₄ 的产量几乎与理论值(图 4C 中的虚线)成正比,且没有出现产量趋于平稳的现象。 我们观察到在培养初期(约第 10 天)有 H₂ 的积累,随后 H₂ 降至检测限以下(< 10 Pa)。这些结果表明,iCH 系统成功实现了低氢分压下产甲烷菌的富集培养。
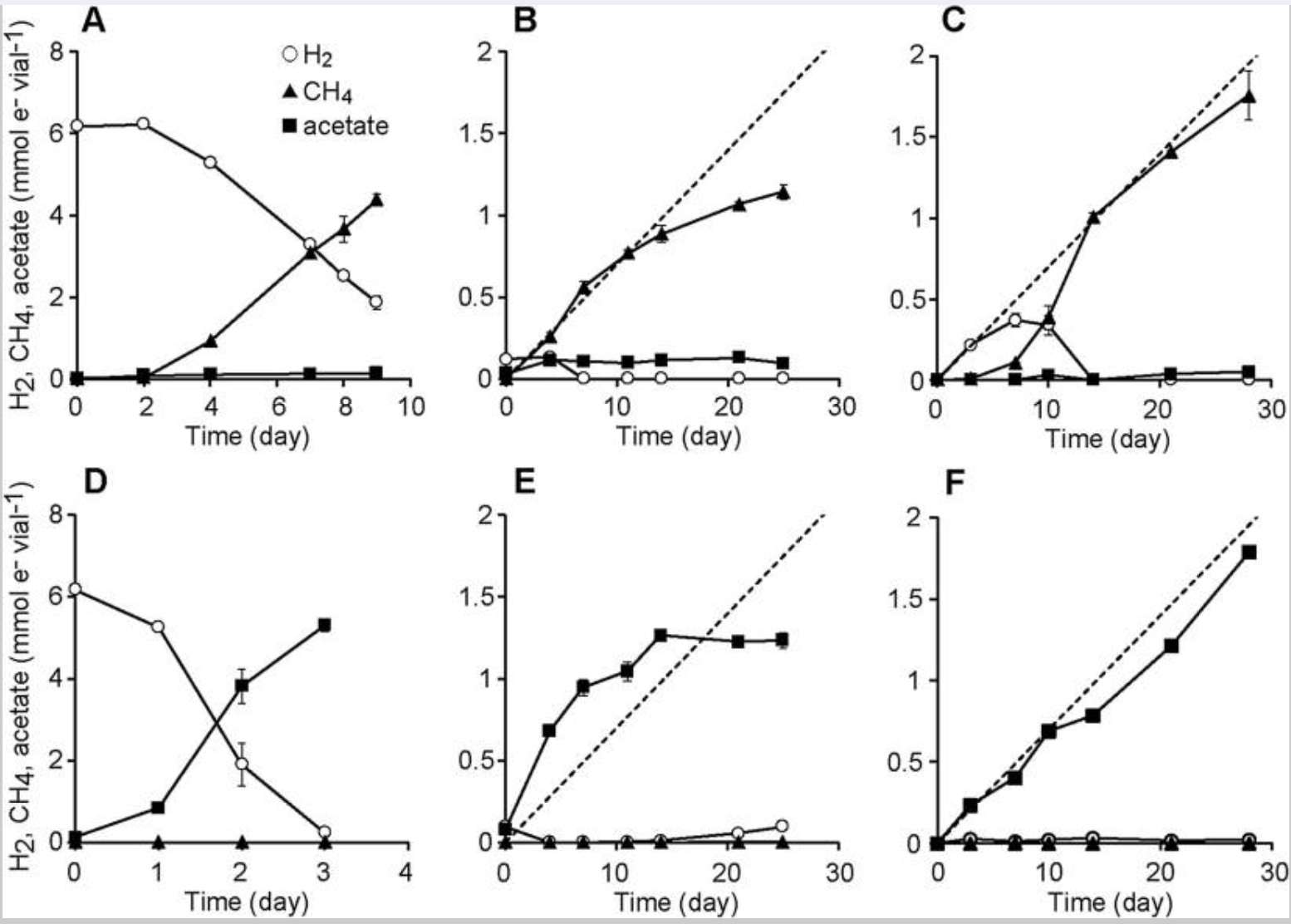
图4:富集培养物中H₂、CH₄和乙酸的生成与消耗
3、微生物群落分析
3.1、产甲烷富集培养物分析
共获取了56,388条16S rRNA基因序列,以97%的序列相似度为阈值将其划分为2949个OTUs。在富集培养物中占优势的所有OTUs(在至少一种富集培养物中相对丰度>3%)在接种土壤中的丰度均很低(约1.2%),这表明利用氢气的微生物得到了充分富集。
在高氢富集组中,两种与Methanobacterium spp.密切相关的OTUs占主导地位(OTU183与Methanobacterium oryzae具有99%的相似度,OTU192与Methanobacterium lacus具有100%的相似度)。相比之下,OTU173和OTU1307(分别与Methanocella arvoryzae和Methanoculleus chikugoensis具有100%的相似度)在iCH培养物中被特异性富集。甲烷胞菌属(Methanocella)和甲烷微菌属(Methanoculleus)经常在包括稻田、泥炭沼泽、海洋和淡水沉积物以及地下环境在内的各种低氢浓度的厌氧环境中被检测为优势氢营养型产甲烷菌。
3.2、产乙酸菌富集培养分析
在高氢富集组中,仅富集出一种与已知产乙酸菌Clostridium magnum密切相关的系统发育型(OTU898)。相比之下,在iCH培养物中选择性地富集出了两种与其他产乙酸菌密切相关的系统发育型(OTU2305和OTU2514,分别与Acetobacterium carbinolicum和Sporomusa sphaeroides具有100%的相似度)。
在Fe⁰富集组中,除了在其他富集组中也检测到的OTU898(C. magnum)和OTU2514(S. sphaeroides)外,占主导地位的还包括与Clostridium glycolicum和Romboutsia lituseburensis密切相关的系统发育型(分别为OTU316和OTU1895)。Clostridium glycolicum是一种著名的产乙酸菌。
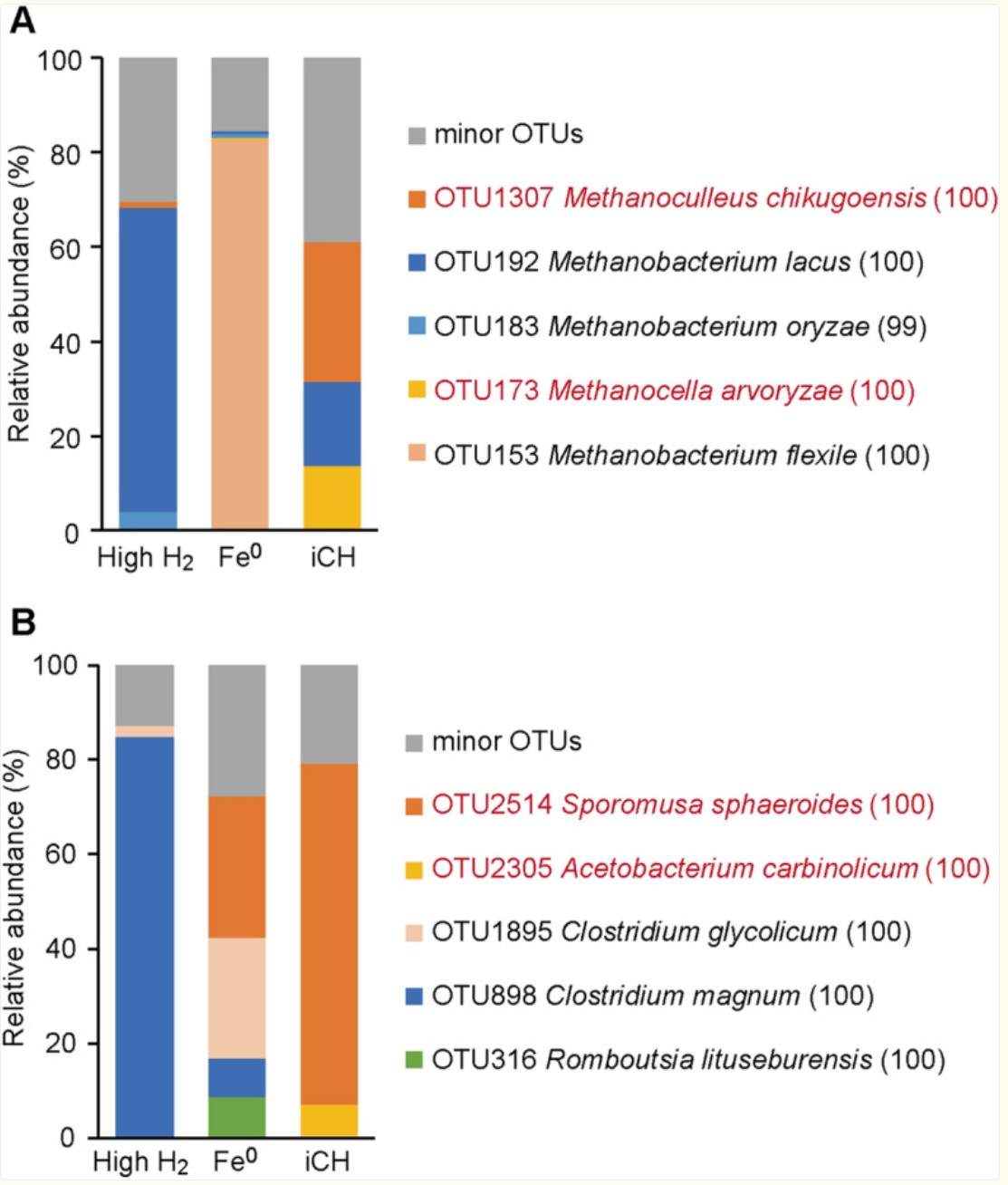
图5:产甲烷菌(A)和产乙酸菌(B)富集培养物中回收的 OTUs 的相对丰度
三、结论
我们利用铁腐蚀反应作为氢源,构建了一个在低氢分压下培养厌氧微生物的简单系统。我们将该系统命名为iCH系统,它可以连续数月供应氢气,并且可以通过改变Fe⁰颗粒的用量和粒径来控制氢气的供应速率。我们证明了iCH系统能够选择性地富集适应低氢分压的厌氧微生物。
尽管本研究仅关注了产甲烷菌和产乙酸菌,但iCH系统同样适用于硝酸盐还原菌、铁还原菌和硫酸盐还原菌等其他利用氢气的微生物的培养。iCH系统还可应用于基于琼脂固化培养基的菌落分离(例如滚管法),这也是我们课题组正在进行的研究。这种培养方法应能实现从地下环境、泥炭土和深海沉积物等低氢可利用性的厌氧环境中,选择性地富集和分离适应甚至特化于低氢分压的未鉴定微生物,这将有助于揭示厌氧环境中氢营养型微生物的新型生态生理学特征。
参考文献
Schink B. Energetics of syntrophic cooperation in methanogenic degradation. Microbiol. Mol. Biol. Rev. 1997;61:262–280. doi: 10.1128/.61.2.262-280.1997.
McInerney MJ, Sieber JR, Gunsalus RP. Syntrophy in anaerobic global carbon cycles. Curr. Opin. Biotechnol. 2009;20:623–632. doi: 10.1016/j.copbio.2009.10.001.
Kato S, Watanabe K. Ecological and evolutionary interactions in syntrophic methanogenic consortia. Microbes Environ. 2010;25:145–151. doi: 10.1264/jsme2.ME10122.
相关产品
HZB428397:甲酸甲烷杆菌DSM 1535 | Methanobacterium formicicum (MF)
HZB501745:康氏甲烷菌DSM 24694 | Methanocella conradii
HZB443012:球形孢子菌DSM 2875 | Sporomusa sphaeroides
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-06-29
编制人:大刘
审稿人:叶凡