肝螺杆菌 putA 突变株在宿主定植和氧化应激中的特性研究
来源:武汉市灰藻生物科技有限公司 浏览量:328 发布时间:2026-05-11 22:22:15
摘要
肝螺旋杆菌是一种革兰氏阴性、螺旋形的微需氧细菌,与易感小鼠株的慢性肠道感染相关,导致肝炎以及结肠癌和肝癌。在密切相关的人类病原体幽门螺旋杆菌中,l-脯氨酸是首选呼吸底物,且在感染患者的胃液中含量显著较高。 此前对幽门螺杆菌和肝螺旋体中脯氨酸分解代谢PutA黄素酶的研究显示,螺旋杆菌PutA在脯氨酸氧化过程中通过将还原黄素电子转移到分子氧中产生活性氧。
我们进一步探讨了脯氨酸作为呼吸底物的偏好,以及通过破坏肝螺旋杆菌中putA基因,脯氨酸代谢对宿主感染期间氧化还原环境的潜在影响。通过氧化应激分析和小鼠感染研究, 最终产生的putA敲除突变株被表征。肝型的putA突变株表现出脯氨酸水平和抗氧化应激能力,相较野生型株,这与脯氨酸作为抗氧化剂的作用相符。
抗应激能力的显著提升归因于脯氨酸含量的增加, 因为putA突变株未观察到抗氧化基因上调。野生型和putA突变肝亨利杆菌株在小鼠中表现出相似感染水平,但在感染putA突变株的小鼠中,炎症显著减少,暗示脯氨酸代谢在体内肝亨利杆菌致病性中起作用。
肝螺杆菌最初是从患有高发性肝炎和肝脏肿瘤的小鼠肝脏组织中分离出来的。随后,发现肝螺杆菌是小鼠肝脏和肠道炎症的致病原,导致肝脏和肠道肿瘤的发生。尽管它经常从肝脏中分离出来并与肝炎有关, 但与其他肠肝螺杆菌属(EHS)一样,肝螺杆菌主要定植于下消化道,与持续性盲肠炎和结肠炎相关。
研究表明,约59%为研究目的而繁殖的小鼠携带肝螺杆菌,或与其他肠肝螺杆菌属共同携带。 因此,实验室小鼠的螺杆菌感染对生物医学研究的潜在影响是一个重大关切。肝螺杆菌是EHS的原型,因此是理解与EHS相关的肠道和肝脏感染分子基础的极佳候选。
本研究的重点是调查脯氨酸代谢在肝螺杆菌氧化应激和感染中的作用。先前的研究表明,脯氨酸在螺杆菌属定植和持续存在于宿主黏膜表面的能力中起着重要作用。 Nagata等人显示,与未感染者相比,幽门螺杆菌感染者的胃液中氨基酸脯氨酸的水平高出10倍。他们还表明,在培养的幽门螺杆菌细胞中,L-脯氨酸的浓度高于所有其他氨基酸,并且L-脯氨酸与L-丝氨酸一起是首选的呼吸底物。
这些观察结果表明,在肠道环境中,脯氨酸是这种胃肠道病原体的关键能量来源。使用签名标签诱变技术,已证明高亲和力脯氨酸特异性转运蛋白基因putP是幽门螺杆菌胃部定植绝对必需的47个基因之一。 也有证据表明,在幽门螺杆菌感染过程中,人血清脯氨酰肽酶会向上调节。
脯氨酰肽酶是一种亚氨基二肽酶,可从甘氨酸-脯氨酸二肽的羧基末端释放脯氨酸或羟脯氨酸,从而提高血清脯氨酸水平。 综合来看,这些观察结果表明脯氨酸代谢在幽门螺杆菌的定植和发病机制中起着重要作用。
革兰氏阴性细菌中的脯氨酸氧化由双功能PutA黄素酶催化,该酶在单个多肽链中含有两个催化结构域。来自肝螺杆菌(PutAHh)和幽门螺杆菌(PutAHp)的PutA共享约64%的氨基酸序列同一性。 脯氨酸氧化的第一步由依赖于黄素腺嘌呤二核苷酸的PRODH结构域执行(图1,反应方案),它将脯氨酸的2e−氧化与细胞质膜的电子传递链系统的还原相偶联。PRODH反应的产物Δ1-吡咯啉-5-羧酸酯(P5C)随后被水解为谷氨酸-γ-半醛(GSA),然后由PutA的依赖NAD+的P5CDH结构域将其氧化为谷氨酸。 通过脯氨酸氧化形成的谷氨酸最终通过α-酮戊二酸进入三羧酸循环。
在先前的一项研究中,PutAHh和PutAHp在与脯氨酸进行催化周转期间被证明会将电子转移给分子氧,形成超氧阴离子和过氧化氢(H2O2)(图1)。大肠杆菌的PutA蛋白在与脯氨酸周转期间几乎没有这种氧反应性。通过大肠杆菌模型系统的氧化应激研究评估了螺杆菌PutA与氧反应的影响,该研究表明PutAHh和PutAHp的酶作用对大肠杆菌有毒。
因此,螺杆菌属中的脯氨酸代谢可能通过PutA形成超氧阴离子以及随后降低脯氨酸水平来改变氧化还原环境,这是合理的,脯氨酸是一种亚氨基酸,已被证明在酵母、植物和哺乳动物细胞中充当强效抗氧化剂。
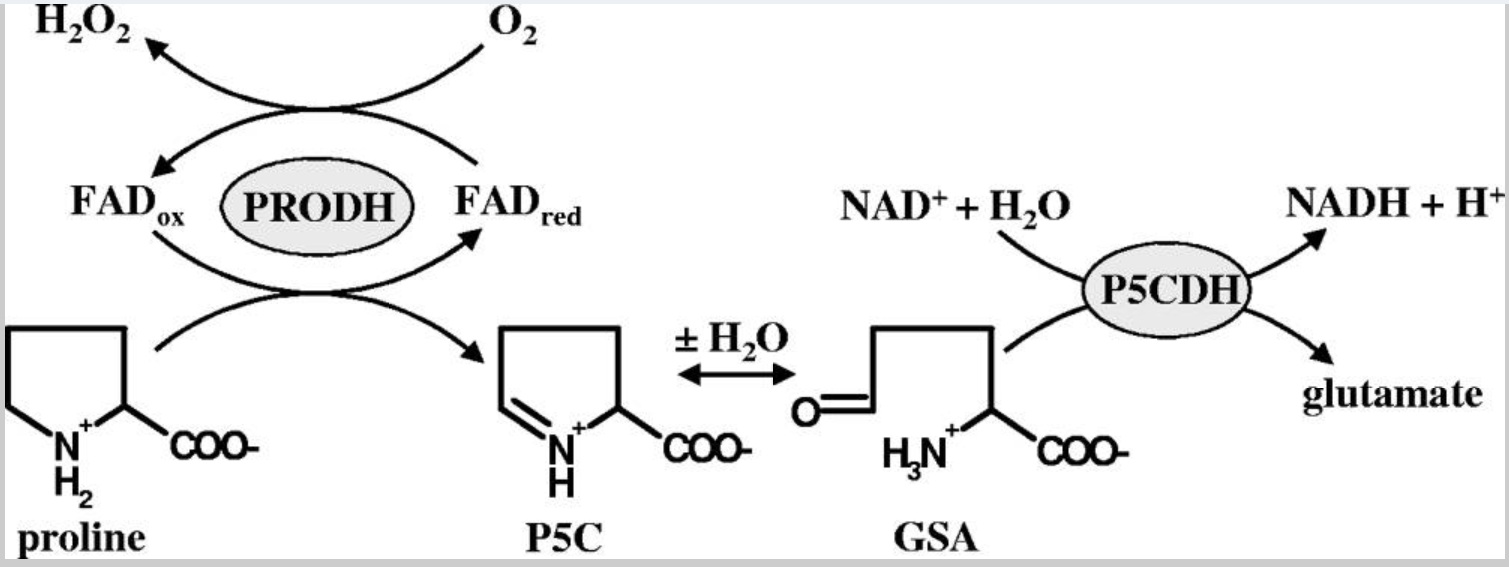
图1:PutA在肝肠杆菌中生成H2O2。图中展示了由PutA的PRODH和P5C脱氢酶(P5CDH)结构域催化的反应。在脯氨酸氧化过程中,PutA的PRODH结构域可以催化还原黄素腺嘌呤二核苷酸电子转移到分子氧,从而形成H2O2。 脯氨酸转化为谷氨酸的整体中间体是P5C和谷氨酸γ半醛(GSA)。。
为了更好地理解脯氨酸代谢在与螺杆菌属感染相关的疾病中的作用,通过盒式诱变法构建了肝螺杆菌的putA突变株。将putA基因敲除株与野生型肝螺杆菌株进行比较,以评估脯氨酸代谢在氧化应激和感染中的作用。 本研究表明,肝螺杆菌中的脯氨酸代谢可以在体外改变氧化还原环境,并影响体内的致病性。
一、材料与方法
1、化学试剂、细菌菌株及培养条件
所有化学试剂和缓冲液,除非另有说明,均购自Fisher Scientific公司和Sigma-Aldrich公司。限制性内切酶和T4 DNA连接酶分别购自Fermentas公司和Promega公司。
克隆和遗传操作使用肝螺杆菌(H. hepaticus)ATCC 51449菌株和大肠杆菌(Escherichia coli)DH5α菌株(BRL)进行。
肝螺杆菌在补充了10%脱纤维羊血(BA)且不含任何抗生素的布鲁氏菌琼脂平板(Difco)上培养。
肝螺杆菌putA突变株在补充了卡那霉素(50 μg/ml)的BA平板上培养。
野生型和putA突变株均在37°C的微需氧环境(5% CO2和1% O2)中培养。
大肠杆菌在含有氨苄青霉素(100 μg/ml)或氯霉素(34 μg/ml)的Luria-Bertani培养基中于37°C有氧培养。
为了分析粪便和组织样本中的肝螺杆菌数量,将样品涂布在含有两性霉素B(10 μg/ml)、万古霉素(10 μg/ml)和头孢哌酮(20 μg/ml)的BA培养基上。
2、通过插入诱变构建肝肠杆菌的突变株
使用肝螺杆菌 ATCC 51449 基因组 DNA 作为模板和 putA 特异性引物(见补充材料中的表 S1)扩增 putA 基因。
根据制造商的建议,将 PCR 片段连接至 pGEM-T 载体(Promega),构建 pGEM-T::putA 质粒。
随后,利用通过 QuikChange(Stratagene)定点诱变技术分别在 745 bp(AflII 引物)和 2728 bp(XhoI 引物)处引入到 putA 基因内的独特 AflII 和 XhoI 位点,将一个卡那霉素抗性盒(1.1 kb)插入到 putA 基因(3.505 kb)中。
将所得的含有卡那霉素插入片段的构建体(pGEM-T::putA::Kan)通过电转化(2.5 kV 脉冲;Gene Pulser;Bio-Rad)导入肝螺杆菌 ATCC 51449T 中。
然后将转化子涂布在含卡那霉素的 BA 平板上,以分离 putA 突变株。
使用上述 putA 特异性引物通过 PCR 确认卡那霉素盒已插入肝螺杆菌的 putA 基因中。
PCR 反应条件为:95°C 30 秒,52°C 1 分钟,72°C 4 分钟,共 30 个循环。
3、野生型与 putA 敲除突变株的生长情况
通过在微需氧条件下培养于 BA 平板上的野生型和 putA 突变肝螺杆菌菌株的对比,评估了破坏 putA 基因及脯氨酸分解代谢对细胞生长、脯氨酸含量以及抗氧化基因表达的影响。
将在 BA 平板上生长的野生型和 putA 突变肝螺杆菌菌株悬浮并稀释于布鲁氏菌肉汤培养基中至约 1.0 × 10⁹ 个细胞/毫升,此处假设 600 nm 处的光密度(OD₆₀₀)为 1.0 时相当于约 1.6 × 10⁹ 个细胞/毫升,如先前所述 。
随后将 3.0 × 10⁸ 个细胞涂布于新鲜的 BA 平板上,分别培养 0、24、48、96、144 和 192 小时。
0 小时的细胞数量为 3.0 × 10⁸ 个细胞/毫升。在 24 至 192 小时期间,通过在不同时间点将 BA 平板上的细胞重新悬浮于布鲁氏菌肉汤培养基中至终体积 1 ml,并根据 OD₆₀₀ 为 1.0 相当于约 1.6 × 10⁹ 个细胞/毫升来估算细胞数量。
所得细胞悬液用于测定脯氨酸含量,并按下文所述分析 PutA、超氧化物歧化酶(SOD)和过氧化氢酶的表达。0 小时的测量是在细胞涂布于 BA 平板之前对细胞悬液进行的。
4、野生型与 putA 突变株中脯氨酸的定量分析
使用上述细胞悬液测定生长 0、24、48、96、144 和 192 小时时的脯氨酸含量。
将每个时间点约 3.0 × 10⁸ 个细胞通过离心沉淀,重悬于 0.5 ml 无菌水中,并煮沸 10 分钟进行裂解。
脯氨酸的含量通过 PRODH 活性测定法进行定量,该方法使用了大肠杆菌 PRODH 结构域构建体 EcPutA86-630,其纯化方法如先前所述 。
简而言之,脯氨酸:二氯酚靛酚氧化还原酶测定在 50 mM Tris 缓冲液(pH 7.5)中进行,方法如先前所述,不同之处在于使用了 EcPutA86-630(100 μg)和 100 μl 细胞提取物。
利用 EcPutA86-630 在测定中 0 至 0.5 mM 脯氨酸范围内的 PRODH 活性标准曲线,来确定总细胞裂解物中的脯氨酸含量。
5、Western 印迹分析
针对纯化的重组 PutAHh 的抗血清由 Proteintech Inc.制备。
重组 PutAHh 的纯化方法如先前所述 (26)。除非另有说明,将约 3 × 10⁸ 个细胞在含有 0.2% Triton 的冰预冷磷酸盐缓冲液(PBS)中通过温和超声破碎 1 分钟(脉冲开启 15 秒,脉冲关闭 30 秒),并将裂解物在 4°C 下以 16,000 × g 离心 10 分钟。
使用二辛可宁酸(Pierce)并以牛血清白蛋白作为标准品,测定细胞裂解物可溶部分的总蛋白浓度。
取 20 μg 或 50 μg 总蛋白样品进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳分离,并按先前所述电转移至聚偏二氟乙烯免疫印迹膜上(孔径 0.2 μm;Bio-Rad)。
然后用 5% 脱脂奶粉封闭聚偏二氟乙烯膜,并与 PutAHh 抗血清(1:20,000 稀释)孵育。
使用单克隆抗 Flag-异硫氰酸荧光素结合物(Sigma-Aldrich)检测免疫反应条带,并使用 LI-COR Odyssey 成像仪显影,方法如先前所述。
PutAHh 抗血清无法识别来自其他细菌来源(如大肠杆菌)的 PutA 酶。
6、氧化应激研究
通过使用三种不同的细胞毒性试剂诱导氧化应激,并通过纸片法进行评估。使用相同的方法,利用 1 M 脯氨酸研究了脯氨酸介导的应激。
将直径为 7.5 mm 的无菌滤纸片置于 BA 平板(100 × 15 mm)上,纸片含有 10 μl 的 1 M 过氧化氢(H₂O₂)、0.2 M 叔丁基过氧化氢(tBH)和 50 mM 百草枯, 这些平板预先划线接种了 putA 突变株或野生型细胞以获得融合生长。
然后将平板置于 2% O₂ 培养箱中。孵育 48 小时后,测量纸片周围的透明区或抑菌圈。 抑菌圈代表从纸片边缘到生长开始处的距离。仅含 H₂O 的纸片未观察到生长抑制。
野生型和 putA 突变肝螺杆菌菌株在每种氧化剂下的平均抑菌圈直径数据来自三次独立实验(每次实验六个样本)。
使用 Student's t 检验(双尾)评估数据的统计学显著性。亲本株与突变株之间观察到的差异在 P 值 小于0.05 时被认为具有统计学显著性。
7、RNA 提取与 RT-PCR
使用在微需氧条件下于 BA 平板上生长的细胞,分析了野生型和 putA 突变肝螺杆菌菌株中 putA、中性粒细胞激活蛋白(napA)和脯氨酸转运蛋白(putP)基因的表达。
按上述描述,使用在 BA 平板上生长 0、24、48、96、144 和 192 小时收获的约 3 × 10⁸ 个细胞,评估了野生型和 putA 突变肝螺杆菌菌株中超氧化物歧化酶(sodF)、 过氧化氢酶(katA)、烷基过氧化氢还原酶 TsaA(tsaA)和硫醇过氧化物酶(tpx)的表达谱。
对于 RNA 提取,细胞在 50 mM Tris-HCl(pH 8.0)、1 mM EDTA 和 50 mM NaCl 中裂解, 并补充 1.5% 十二烷基硫酸钠,在 95°C 下处理 5 分钟。总 RNA 经酚抽提和沉淀,然后溶解于焦碳酸二乙酯处理过的水中。 然后在 260 nm 处测定分离的 RNA 浓度。
将收集的 RNA 逆转录以合成总 cDNA,反应体系总体积为 20 μl,包含每种脱氧核糖核苷三磷酸 1 mM、25 U 鼠白血病病毒(MuLV)逆转录酶(RT)、1 U RNase 抑制剂和 2.5 μM 随机六聚体。
所有试剂均来自 Invitrogen。 反应在 MJ mini-thermal cycler 装置(Bio-Rad)中于 42°C 进行 30 分钟,随后在 95°C 下进行 10 分钟步骤以使酶变性,然后冷却至 4°C。所得 cDNA 随后用作 PCR 扩增的模板。致死性膨胀毒素(CdtB)基因产物(编码 cdtB 基因的亚基 B)用作上样对照。
用于 PCR 扩增的引物列于补充材料的表 S1 中。PCR 条件为:95°C 30 秒,52°C 1 分钟,72°C 2 分钟,共 30 个循环。为了分析 sodF、katA、tsaA 和 tpx,反应在 20 个循环时终止,以确保扩增仍处于指数期。 PCR 产物通过溴化乙锭染色显色,并使用 Bio-Rad Gel Doc 系统成像。
8、小鼠感染
所有动物实验均按照 NIH 关于动物妥善护理和处理指南进行。小鼠在内布拉斯加大学林肯分校的动物研究设施中饲养和维护。
从无特定病原体的 Jackson Laboratory(Bar Harbor, ME)购买了 6 周龄的无螺杆菌认证的近交系雄性 A/J 小鼠。
在整个研究期间,动物喂食高压灭菌的小鼠饲料,并提供水和垫料,换笼操作在层流罩下进行。动物以每五个为一组饲养在微隔离笼中,光照和黑暗周期各为 12 小时。
肝螺杆菌参考菌株 3B1/Hh-1(ATCC 51449T)在补充了 5%(体积/体积)羊血(TSA-BA;Remel, Lenexa, KS)的胰蛋白酶大豆琼脂上繁殖,并在微需氧条件下(Mitsubishi Gas Chemical Co. Inc., New York, NY)于 37°C 孵育。
参考株和 putA 突变株在 TSA-BA 平板上生长,并在 PBS(pH 7.2)中洗涤后通过离心收获。接种物的 OD₆₀₀ 调整为 0.32(约 5 × 10⁸ 个细胞/毫升)。
十只小鼠腹腔注射 0.5 ml 野生型或 putA 突变细胞的细菌悬液。十只对照小鼠假接种 0.5 ml 无菌 PBS。
在第 1 周和第 3 周后从笼中采集粪便颗粒样本。接种后第 21 天,通过一氧化碳过量吸入对小鼠实施安乐死。
9、肝脏、盲肠和粪便标本的定性分析
将接种后第 21 天从笼中采集的约 200 mg 粪便颗粒在 5 ml 无菌 PBS 中匀浆。将悬液(200 μl)涂布于含有两性霉素 B(10 μg/ml)、万古霉素(10 μg/ml)和头孢哌酮(20 μg/ml)的 TSA-BA 平板上。平板在 37°C 微需氧环境(1% O₂,5% CO₂)中孵育 72 至 96 小时。
在接种后第 21 天对实施安乐死的小鼠的组织进行收获,用于定性、定量和组织病理学分析。
使用组织捣碎机(Biospec Products, Inc.)将肝脏(左叶、右叶、尾状叶和部分中叶)和盲肠(取出一部分用于组织病理学检查后的全部组织)切片匀浆,并将每种组织悬液的 200 μl 涂布于含抗生素的 TSA-BA 平板上,以确定肝螺杆菌的存在。 如下文所述,还通过实时 PCR 评估组织和悬液中肝螺杆菌的存在。
10、肝脏和盲肠的定量分析
由于肝螺杆菌呈连续扩散的菌苔生长,无法计数细菌菌落。相反,使用实时 PCR 估算不同组织中肝螺杆菌的数量,方法如先前所述。
使用 DNeasy 组织试剂盒(Qiagen)从 25 mg 匀浆的肝脏和盲肠组织中提取 DNA。 使用 Promega 的 Plexor 引物设计工具设计的 6-羧基荧光素标记的肝螺杆菌特异性 cdtB 引物(见补充材料中的表 S1)进行实时 PCR(iCycler Thermal Cycler)。
针对已知基因组进行 BLAST 搜索, 验证 cdtB 引物和 cdtB 基因探针(编码致死性膨胀毒素的亚基 B)对肝螺杆菌具有高度特异性。使用含有 12.5 μl Supermix(Promega)、正向和反向引物各 200 nM 浓度以及 5 μl 组织 DNA(相当于 1.0 mg 组织)的 25 μl 混合物进行实时 PCR 分析。
实时 PCR 条件为:95°C 1 个循环 3 分钟,然后 95°C 30 秒和 65°C 30 秒,共 40 个循环。 实时 PCR 产生 110 bp 的扩增产物。标准曲线(未显示)由 10² 至 10⁷ fg 范围的肝螺杆菌基因组 DNA 样品生成,据此估算每个样品的基因组拷贝数,方法如先前所述 。
11、组织病理学研究
由肝脏中叶、盲肠、近端结肠和脾脏组成的组织标本在 10% 缓冲福尔马林中固定过夜,进行处理并包埋于石蜡中。切片(5 μm 厚)用苏木精-伊红染色,并按先前所述对组织病理学变化进行半定量评分 (37)。
门静脉炎症评分标准为:0(无炎症),1(少数门静脉束有轻度浸润),2(几乎所有门静脉束有轻至中度浸润)。小叶炎症和/或肝细胞坏死分级为:0(无),1(轻度),2(中度)。胆道病变和卵圆细胞改变评分标准为:0(无病变),1(偶发或轻度胆管炎)。
使用 SPSS 统计软件,通过非参数 Mann-Whitney 检验分析感染野生型和 putA 突变肝螺杆菌菌株的小鼠组织的组织病理学评分,统计学显著性设定为 P 值 小于0.05。
二、结果
1、putA 基因的破坏
肝螺杆菌中的 putA 基因通过插入卡那霉素盒被破坏。
通过比较 putA 突变株的约 4.6 kb PCR 产物与野生型菌株的约 3.5 kb 产物,确认了该盒的插入。 分别通过 RT-PCR 和 Western 印迹分析在转录和蛋白水平上确认了基因的破坏(图 2)。针对 putA cDNA 的 RT-PCR 未产生任何产物,而野生型菌株则观察到显著产物。
图 2 显示了来自野生型肝螺杆菌的约 3.5 kb 条带,表明全长的 putA 基因有表达。在野生型肝螺杆菌菌株中也观察到额外的较小条带,这很可能是由于 putA cDNA 内的非特异性引物结合事件造成的。 使用针对纯化的全长肝螺杆菌 PutA 制备的抗体进行 Western 印迹分析。
在突变株中未观察到预期大小(133.4 kDa)的免疫反应条带,但在野生型菌株中检测到了对应于 PutAHh 的 133 kDa 条带。 为了确定没有极性效应,通过 RT-PCR 测试了野生型和突变株中邻近的上游 putP 和下游 napA 基因的表达水平。
putP 基因编码高亲和力脯氨酸转运蛋白,而 napA 基因编码中性粒细胞激活蛋白。 两种菌株均显示出相似的 putP 和 napA 基因表达水平,表明这些基因未被 putA 基因的卡那霉素盒诱变所破坏(数据未显示)。
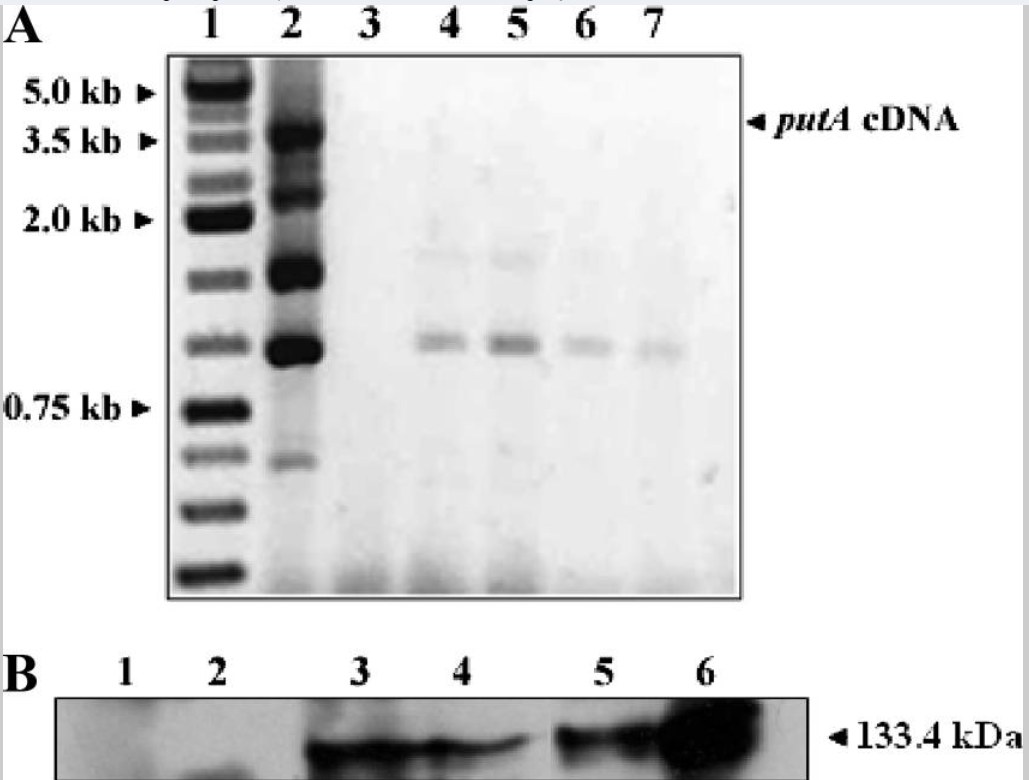
图2:RT-PCR和西方墨迹分析确认了肝肝菌中putA基因的破坏。(A)使用putA基因特异性引物进行RT-PCR。 车道:1条,kb梯;2,野生型肝肝菌株;3,空;4至7个不同群体筛查肝炎的putA缺失。(B)野生型和PutA H. hepaticus菌株蛋白提取物的西方印迹分析。 每个细胞裂解物样本装载了两种不同量的总蛋白。通道:1和2,放置A突变细胞裂解物(分别为20 μg和50 μg);3和4,野生型细胞裂解物(分别为50微克和20微克);5和6, 纯化重组PutAHh(分别为20微克和50微克)。
2、putA 突变株的氧化应激和脯氨酸毒性抗性增强
我们通过使用过氧化氢(1 M)、tBH(0.2 M)和百草枯(0.05 M)进行纸片法实验,表征了野生型和 putA 突变株的体外氧化应激反应。putA 突变株在 H₂O₂作用下表现出的抑菌圈直径比野生型菌株小 2 倍(表 1)。 在 tBH 和百草枯作用下,putA 突变株的抑菌圈直径比野生型菌株小 3 倍(表 1)。
因此,与亲本野生型菌株相比,突变株表现出显著更高的氧化应激抗性。 由于 PutA 已被证明在体外会产生 ROS,我们确定了添加脯氨酸是否也会导致应激和细胞死亡。在测试了广泛的脯氨酸浓度范围后,在这些实验的滤纸片上使用了 1 M 脯氨酸。
野生型菌株对脯氨酸表现出高度敏感性,其抑菌圈直径与 H₂O₂应激下观察到的相似。相比之下,脯氨酸对 putA 突变株没有任何影响,因为未观察到抑菌圈。
这些观察结果与我们之前的结果一致, 即在大肠杆菌中过表达 PutAHh 时,补充脯氨酸(5 mM)被证明对细胞有毒性 (26)。此处表明,当脯氨酸水平升高时,PutAHh 的酶促作用对肝螺杆菌也有毒性。
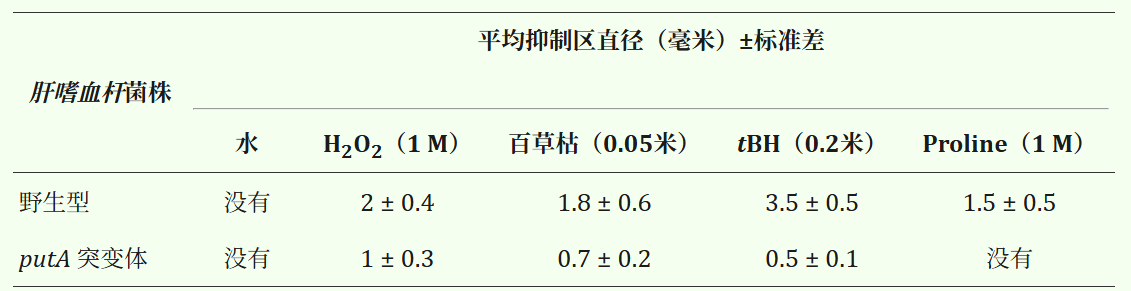
表1:通过处理10微升所示化合物的滤纸片测量抑制区。水作为对照,未形成生长抑制区。结果是三个独立实验±标准差(SD)的平均值。 根据Student's t检验,具有H2O2、百草枯和tBH的putA突变株抑制区显著低于亲本野生型株(基于Student's t检验)的95%置信水平(P < 0.05)。
3、putA 突变株中脯氨酸水平升高
细胞在 BA 平板上培养长达 192 小时。野生型和 putA 突变株的生长速率之间未观察到显著差异(图 3A)。在生长期间的不同时间点测量了细胞中的脯氨酸含量。在正常生长条件下,突变株的细胞内脯氨酸水平高于野生型菌株(图 3B)。在整个 192 小时的生长期间,突变株的脯氨酸水平约为野生型菌株的 2 倍。
这一结果与突变株在缺乏 PutA 的情况下无法分解代谢脯氨酸的能力缺失相符。 由于脯氨酸被认为是一种具有自由基清除特性的抗氧化剂 (1),putA 突变株中较高的脯氨酸水平可能解释了在突变株中观察到的应激抗性增强,以及缺乏 PutA 产生的 ROS。
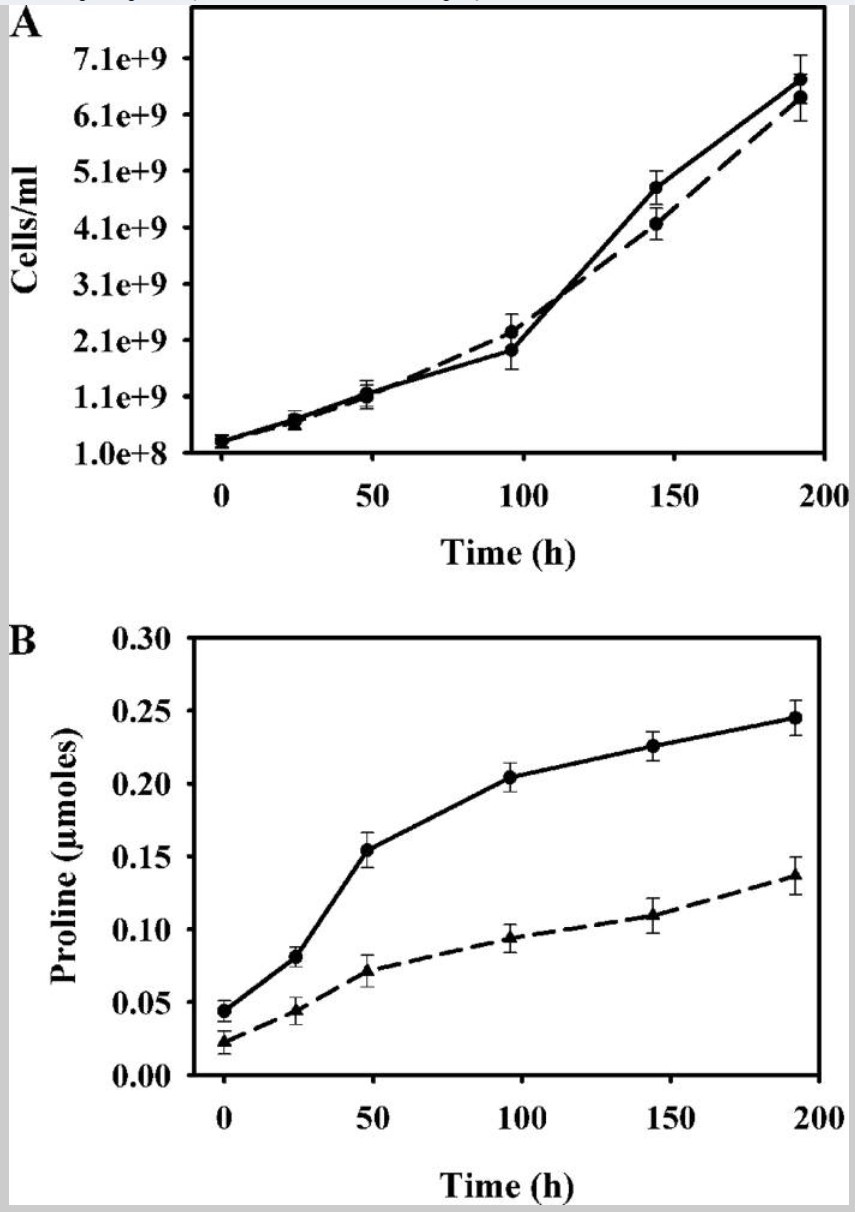
图3:(A)野生型(实心曲线)和putA突变体(虚线曲线)肝氏菌株的生长曲线,在BA板上孵育时间为0、24、48、96、144和192小时。细胞在每个时间点重新悬浮于布鲁氏菌培养基中进行OD测量。 (B)野生型的脯氨酸含量,并将A突变肝菌株在生长0、24、48、96、144和192小时的BA板上。每个时间点约使用3×108细胞测定脯氨酸含量。
三、讨论
研究显示,感染幽门螺旋桨杆菌患者的肠道中脯氨酸水平较高,且在不同研究中,脯氨酸补充被观察到对幽门螺旋菌增殖至关重要。这些有趣的观察促使我们对幽门螺旋杆菌和肝扁桃蛋白的PutA蛋白进行了表征。
我们发现,类似于人类PRODH,螺旋杆菌属的PutA在与脯氨酸和分子氧的交替过程中产生超氧化物阴离子。PutAHh和PutAHp的氧化酶活性被认为不寻常,因为其他革兰氏阴性菌的PutA同源蛋白表现出极低的氧化酶活性。 PutAHh和PutAHp的氧化酶活性对大肠杆菌有毒,严重抑制细胞存活率。
因此,除了脯氨酸作为能量底物的作用外,脯氨酸代谢可能对肝氏的氧化还原环境产生双重影响,PutA活性产生ROS,脯氨酸积累则保护其免受氧化应激。 因此,测试PutA和脯氨酸代谢在肝肝杆菌中的病理生理作用具有重要性。
为探讨脯氨酸代谢对肝螺旋杆菌生理和感染的影响,产生了肝螺旋杆菌的putA突变株。突变株被发现能积累更多的脯氨酸,且对氧化应激的抵抗力显著优于野生型株。 突变株能够在高浓度氧化剂下存活,表明PutA的缺失会导致氧化还原环境发生有利于细胞生存的变化。
putA突变株抗氧化应激性增强的主要原因可能是脯氨酸具有羟基自由基清除特性。 或者,putA突变株中抗氧化酶的表达水平可能更高。然而,我们发现相反趋势,野生型株表现出更高的SOD和过氧化氢酶表达水平,而putA突变株则高于PutA突变株。因此,脯氨酸似乎在肝肝杆菌中具有细胞保护作用, 这一作用此前尚未在该病原体中被证实。
高浓度脯氨酸还被证明对肝型H. hepaticus野生型株有毒,但对putA突变株无致命影响。这些结果证明PutA活性会导致肝肝型菌株的脯氨酸毒性和细胞死亡。
PutA的毒性效应可能由ROS形成或中间体P5C的产生介导,后者已被证明能诱导哺乳动物细胞凋亡。正如之前一项研究报道的,螺旋杆菌很可能存在于脯氨酸水平较高的生态位; 因此,它必须在脯氨酸代谢的有益特性与PutA活性的有害促氧化作用之间取得平衡。
野生型细胞中sodF和katA基因表达相较于putA突变株在正常生长条件下的意外增加,表明肝螺旋杆菌利用SOD和过氧化氢酶来对抗PutA活性的不良反应。 由于早期关于幽门螺杆菌的研究显示脯氨酸是重要的呼吸底物,肝螺旋杆菌可能利用SOD和过氧化氢酶控制PutA活性的毒性副作用,同时受益于脯氨酸代谢的能量和生长特性。
组织样本分析显示,接种肝肠杆菌野生型小鼠与接种putA突变株的小鼠肝脏和盲肠定殖率无显著差异,表明putA基因的破坏不会影响肝肝菌的定植能力。 然而,感染putA突变株的小鼠肝组织组织病理学检查显示,炎症明显低于接种野生型株的小鼠肝样本。
这表明,虽然PutA对宿主定殖可能不关键,但PutA和脯氨酸代谢有助于促进肝肝菌的发病机制。 肝脏样本观察到的炎症主要由肝脏嗜血杆菌感染部位周围的中性粒细胞活化和ROS生成引起。显然,缺乏PutA活性和脯氨酸的利用减少了ROS引起的组织损伤,甚至可能减少宿主中性粒细胞的激活。
肝肝H. A突变体中脯氨酸的积累可能导致ROS水平降低,从而减少或减缓炎症反应。这与putA突变株抗氧化应激性增强相符。相反,野生型株中的PutA活性有助于加速或加重炎症和肝螺旋杆菌感染的发病机制。
由于脯氨酸作为ROS清除剂和通过PutA促进氧化剂的双重作用,脯氨酸代谢非常适合调节肝螺旋杆菌感染的炎症反应和致癌机制。 未来的研究将重点揭示脯氨酸代谢是否会调用参与氧化应激反应和炎症的某些信号通路。例如,酸盐酸酶和铁结合蛋白NapA等毒力因子在幽门螺杆菌中通过氧化应激被上调, 以帮助保护幽门螺杆菌并使慢性炎症得以持续(5, 9, 21)。
野生型肝螺旋杆菌株的过氧化氢酶表达水平高于putA突变株,表明通过PutA进行脯氨酸氧化在细胞内产生氧化还原信号,帮助维持过氧化氢酶表达,甚至可能产生毒力。 还需进行更多实验以确定破坏putA基因是否会减弱其他潜在毒力因子的表达。还将探讨脯氨酸运输,以区分脯氨酸的细胞内和细胞外储藏。
研究显示,脯氨酸摄取在人类病原体金黄色葡萄球菌的存活中起着关键作用, 因为编码高亲和力脯氨酸转运蛋白的putP基因缺失会导致多种感染模型中显著减弱。
参考文献
1、Alia, P. Mohanty, and J. Matysik. 2001. Effect of proline on the production of singlet oxygen. Amino Acids 21195-200.
2、Aslan, M., Y. Nazligul, M. Horoz, C. Bolukbas, F. F. Bolukbas, N. Aksoy, H. Celik, and O. Erel. 2007. Serum prolidase activity and oxidative status in Helicobacter pylori infection. Clin. Biochem. 4037-40.
3、Bayer, A. S., S. N. Coulter, C. K. Stover, and W. R. Schwan. 1999. Impact of the high-affinity proline permease gene (putP) on the virulence of Staphylococcus aureus in experimental endocarditis. Infect. Immun. 67740-744.
相关产品
HZB362806:肝螺旋杆菌ATCC 51449 | Helicobacter hepaticus Fox et al.
HZB909279:肝螺旋杆菌CCUG 44777 | Helicobacter hepaticus
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-05-11
编制人:大刘
审稿人:叶凡