从基础开始学习原核细胞05:古菌的细胞壁、原核生物的蛋白质分泌
来源:武汉市灰藻生物科技有限公司 浏览量:203 发布时间:2026-05-11 21:14:42
1、古菌的细胞壁
在被确认为一个独立的生命领域之前,古菌曾被简单地归类为革兰氏阳性或阴性。然而,它们的染色反应与特定的细胞壁结构之间,并不像细菌的革兰氏染色那样有可靠的对应关系。
古菌的细胞壁在结构和化学组成上都与细菌不同。它们的细胞壁不含肽聚糖(peptidoglycan),并且在化学成分上表现出极大的多样性。
许多古菌具有一层厚而均匀的细胞壁,外观类似于革兰氏阳性细菌(图1a)。这类古菌通常呈革兰氏阳性染色。其细胞壁的化学成分因物种而异,但通常由复杂的杂多糖(heteropolysaccharides)构成。
例如,甲烷杆菌属(Methanobacterium)和其他一些产甲烷古菌(methanogens)的细胞壁含有假肽聚糖(pseudomurein)(图2)。
这是一种类似肽聚糖的聚合物,但其交联结构中使用的是L-型氨基酸而非D-型氨基酸;其糖骨架中含有N-乙酰塔罗糖胺糖醛酸(N-acetyltalosaminuronic acid),而非N-乙酰胞壁酸(N-acetylmuramic acid);并且其糖苷键为β(1→3)键,而非β(1→4)键。
其他古菌,如甲烷八叠球菌属(Methanosarcina)和嗜盐的盐球菌属(Halococcus),其细胞壁则含有类似于动物结缔组织中的硫酸软骨素(chondroitin sulfate)的复杂多糖。
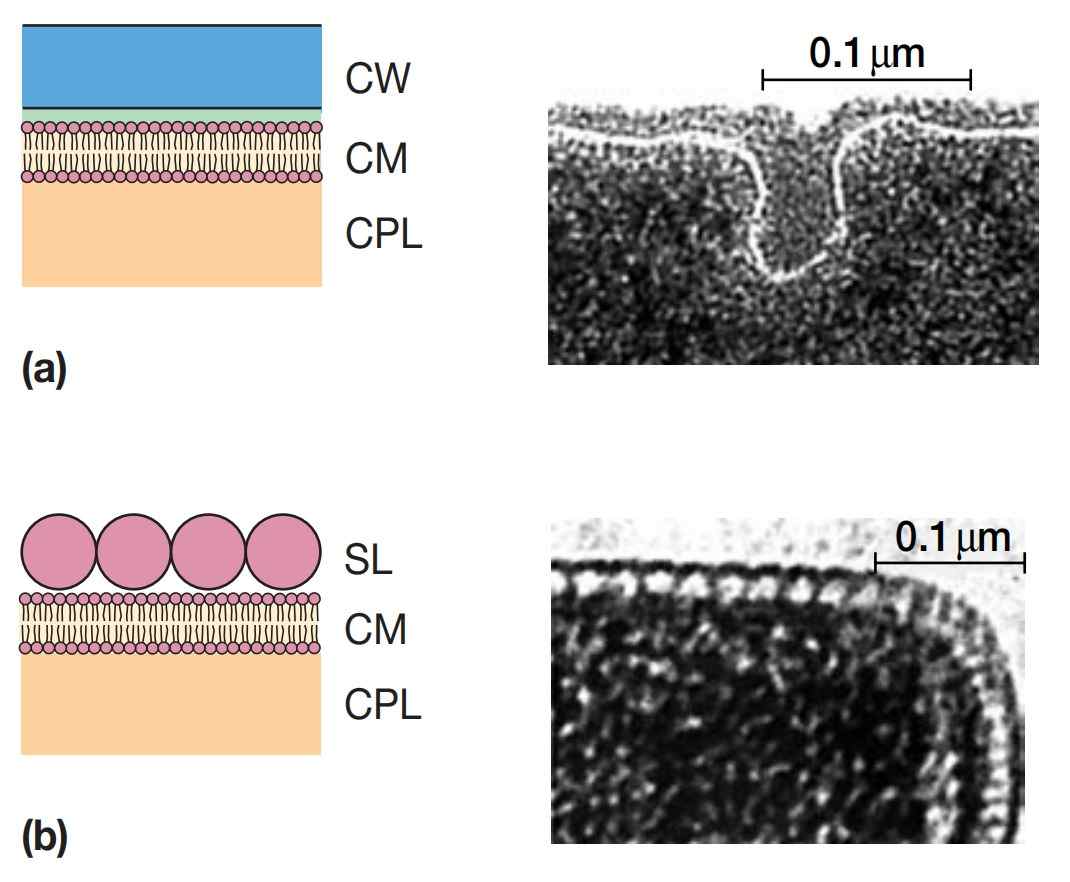
图1 古菌的细胞包膜(Cell Envelopes of Archaea)。示意图及电镜照片:(a)甲酸甲烷杆菌(Methanobacterium formicicum) 和(b)硫化叶菌(Thermoproteus tenax) 。CW:细胞壁(cell wall);SL:表层(surface layer);CM:细胞膜或质膜(cell membrane or plasma membrane);CPL:细胞质(cytoplasm)。
另一些呈革兰氏阴性染色的古菌,在其质膜外有一层糖蛋白(glycoprotein)或蛋白质(protein)(图1b)。这层结构厚度可达20至40纳米。有时甚至存在两层:一层电子致密层,以及包裹在其外的一层鞘。
一些产甲烷菌(如Methanolobus)、嗜盐古菌(如Halobacterium)和极端嗜热菌(如Sulfolobus, Thermoproteus, Pyrodictium)的细胞壁中含有糖蛋白。相比之下,其他产甲烷菌(如Methanococcus, Methanomicrobium, Methanogenium)和极端嗜热菌Desulfurococcus则具有纯蛋白质构成的细胞壁。
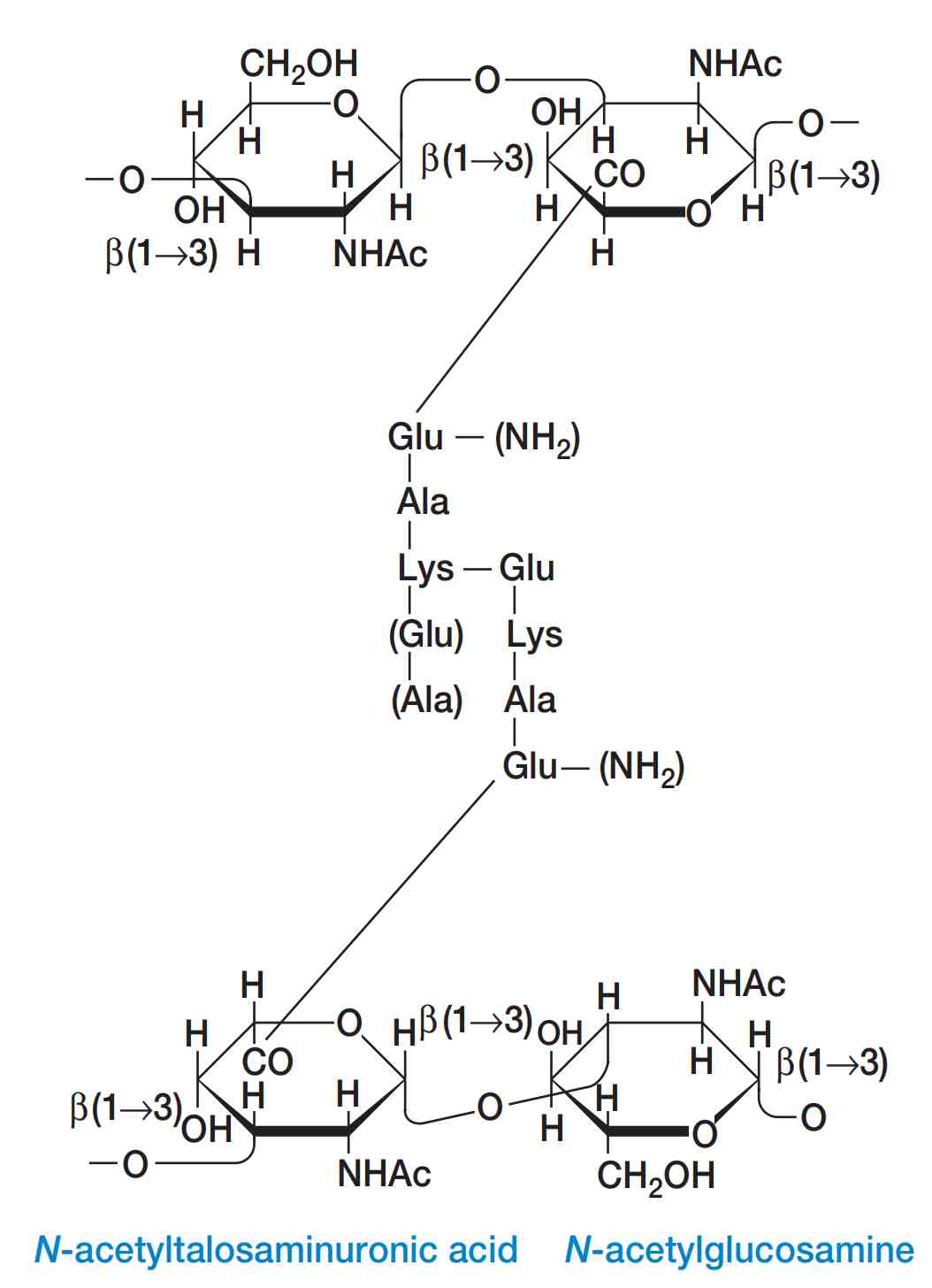
图2 假肽聚糖的结构(The Structure of Pseudomurein)。括号内的氨基酸和氨基并非总是存在。Ac代表乙酰基(acetyl group)。
2、原核生物的蛋白质分泌
原核细胞的包膜(即古菌和革兰氏阳性菌的质膜,以及革兰氏阴性菌的质膜和外膜)对大分子进出细胞构成了显著屏障。
然而,许多重要结构位于细胞壁之外。构成这些结构的大分子是如何被运出细胞进行组装的?此外,原核生物会向周围环境释放胞外酶(exoenzymes)和其他蛋白质。这些蛋白质又是如何穿过细胞包膜的?
显然,原核生物必须具备分泌蛋白质的能力。过去几十年,蛋白质分泌通路的研究迅速发展。这不仅因为其基础重要性,也因为某些分泌机制与病原菌的致病性相关。
对分泌机制的理解还可用于疫苗开发和多种工业过程。由于对古菌的蛋白质分泌系统了解甚少,本次主要概述细菌的蛋白质分泌通路。
(1)细菌蛋白质分泌概述
蛋白质分泌的难度取决于细菌是革兰氏阳性还是阴性。革兰氏阳性菌分泌蛋白质时,只需将蛋白质转运穿过质膜。一旦穿过质膜,蛋白质要么通过相对多孔的肽聚糖层进入外部环境,要么嵌入或附着在肽聚糖上。
革兰氏阴性菌在分泌蛋白质时面临更多障碍。它们同样需要将蛋白质转运穿过质膜,但为了完成分泌,蛋白质还必须能避开周质空间(periplasmic space)中蛋白水解酶的降解,并且要穿过外膜。
在革兰氏阳性和阴性菌中,将蛋白质穿过质膜的主要通路是Sec依赖通路(Sec-dependent pathway)(图3)。在革兰氏阴性菌中,蛋白质可以通过多种不同机制穿过外膜。其中一些机制完全绕过Sec依赖通路,直接将蛋白质从细胞质运送到细胞外(图4)。
此处描述的所有蛋白质分泌通路在某个步骤都需要消耗能量。能量通常由ATP和GTP等高能分子的水解提供,有时质子动力(proton motive force)也起作用。
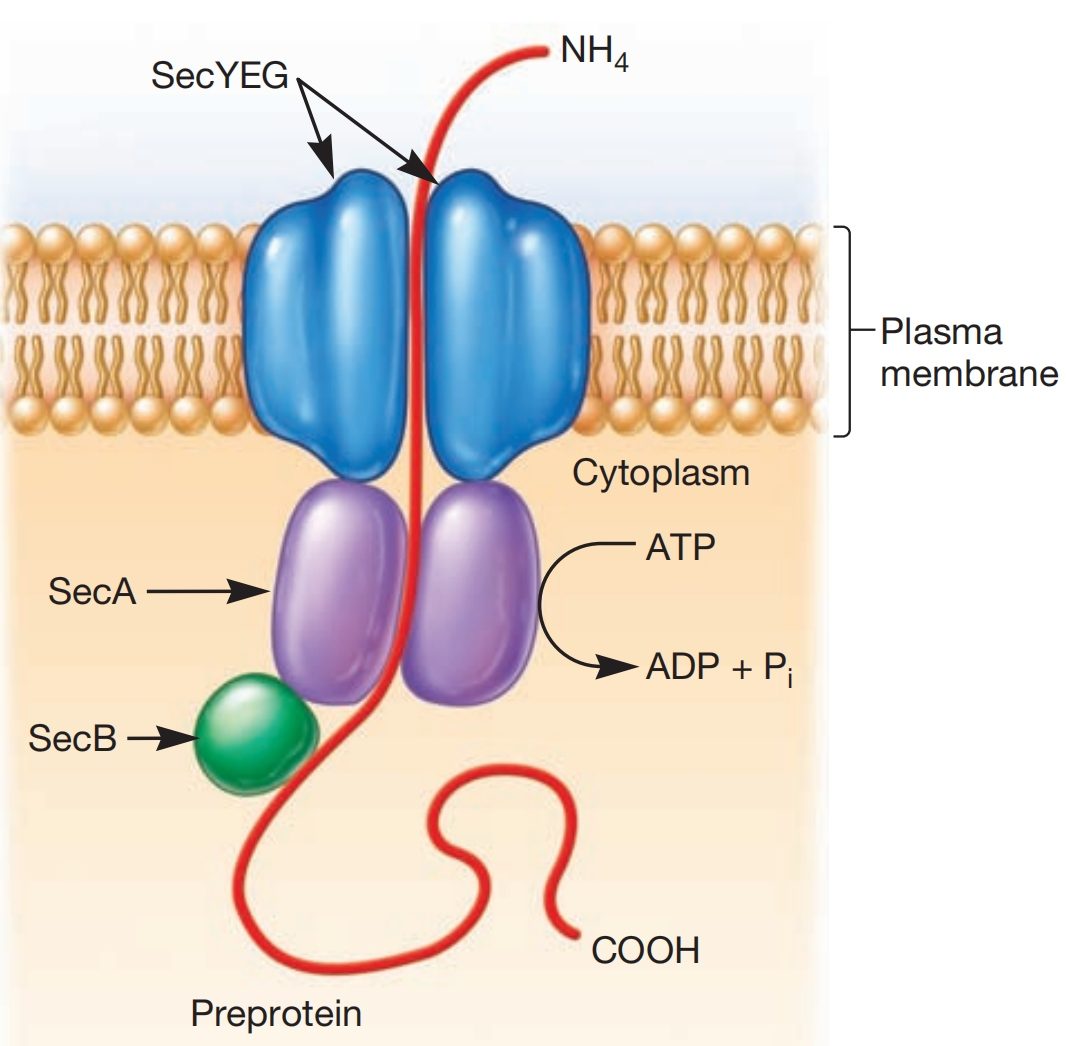
图3 Sec依赖通路(The Sec-Dependent Pathway)。大肠杆菌(E. coli)的Sec依赖通路。
(2)Sec依赖通路
Sec依赖通路,有时也称为一般分泌通路(general secretion pathway),在所有三个生命域中都高度保守(图3)。它能将蛋白质跨质膜转运,或将蛋白质整合到膜中。
通过此通路转运的蛋白质最初被合成为前体蛋白(preproteins)。前体蛋白的氨基末端带有一个信号肽(signal peptide),可被Sec转运机器识别。
信号肽合成后不久,一类名为分子伴侣(chaperone proteins)的特殊蛋白(如SecB)会与之结合。这有助于延迟蛋白质的折叠,从而帮助前体蛋白以适合转运的构象到达Sec转运机器。
有证据表明,蛋白质的转运可以在核糖体完成其合成之前就开始。某些Sec蛋白(SecY、SecE和SecG)被认为在膜上形成一个通道,前体蛋白由此通过。另一种蛋白(SecA)会与SecYEG蛋白和SecB-前体蛋白复合物结合。SecA作为一种马达,利用ATP水解的能量,将前体蛋白(而非分子伴侣)穿过质膜。
当前体蛋白从质膜另一侧出现并脱离分子伴侣后,一种名为信号肽酶(signal peptidase)的酶会切除信号肽。随后,蛋白质折叠成正确的形状,并在必要时形成二硫键。
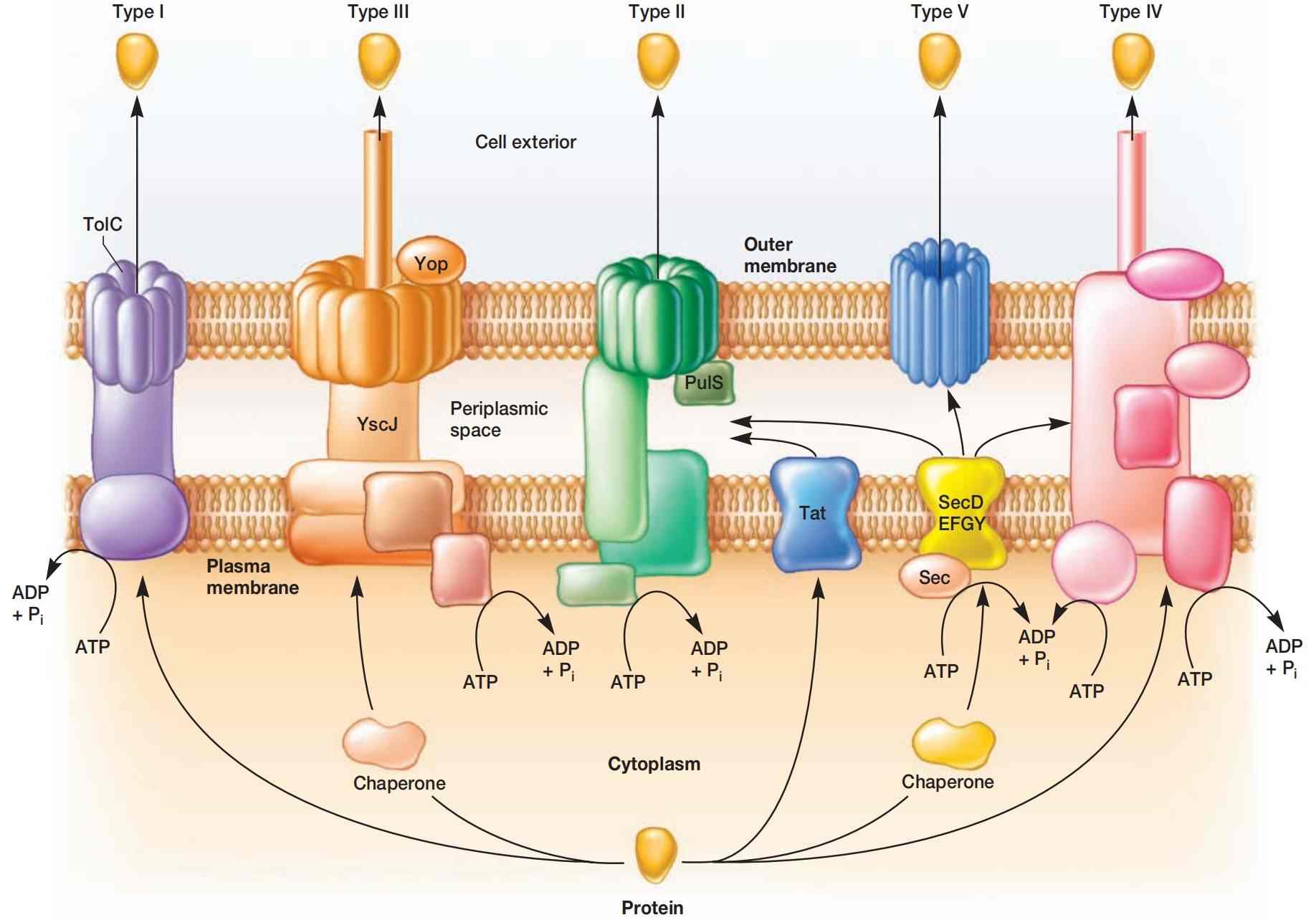
图4 革兰氏阴性菌的蛋白质分泌系统(The Protein Secretion Systems of Gram-Negative Bacteria)。 图中展示了革兰氏阴性菌的五种分泌系统。Sec依赖通路和Tat通路将蛋白质从细胞质运送至周质空间。 II型、V型及部分IV型系统完成由Sec依赖通路启动的分泌过程。 Tat系统仅向II型通路输送蛋白质。 I型和III型系统则绕过Sec依赖和Tat通路,直接将蛋白质从细胞质经外膜运送到胞外。 IV型系统既可与Sec依赖通路协同工作,也可独立将蛋白质转运至胞外。 Sec依赖通路和III型通路转运的蛋白质由分子伴侣蛋白递送至相应系统。
(3)革兰氏阴性菌的蛋白质分泌
目前在革兰氏阴性菌中已鉴定出五种蛋白质分泌通路(图4)。革兰氏阴性菌拥有第二层外膜,蛋白质必须穿过这层膜。
革兰氏阴性菌利用II型和V型通路,将已由Sec依赖通路穿过质膜的蛋白质再转运穿过外膜。I型和III型通路不与Sec系统转运的蛋白质相互作用,因此被称为Sec非依赖通路。IV型通路有时与Sec依赖通路相连,但通常独立运作。
II型蛋白质分泌通路存在于多种动植物病原菌中,如胡萝卜软腐欧文氏菌(Erwinia carotovora)、肺炎克雷伯菌(Klebsiella pneumoniae)、铜绿假单胞菌(Pseudomonas aeruginosa)和霍乱弧菌(Vibrio cholerae)。它负责分泌多种降解酶,如普鲁兰酶(pullulanases)、纤维素酶(cellulases)、果胶酶(pectinases)、蛋白酶(proteases)和脂肪酶(lipases),以及其他蛋白质,如霍乱毒素(cholera toxin)和菌毛蛋白(pili proteins)。
II型系统非常复杂,可包含多达12至14种蛋白质,其中大部分似乎是内在膜蛋白(integral membrane proteins)(图4)。
尽管II型系统的某些组分跨越质膜,但它们似乎只负责将蛋白质转运穿过外膜。在大多数情况下,Sec依赖通路首先将蛋白质转运穿过质膜,然后II型系统完成分泌过程。
在革兰氏阴性和阳性菌中,还存在另一种名为Tat通路(Tat pathway)的质膜转运系统,可将蛋白质转运穿过质膜。在革兰氏阴性菌中,这些蛋白质随后被递送给II型系统。Tat通路与Sec系统不同,它能转运已经折叠好的蛋白质。
V型蛋白质分泌通路是最新发现的分泌系统。它们同样依赖Sec依赖通路将蛋白质穿过质膜。然而,一旦进入周质空间,其中许多蛋白质能够在外膜上自行形成通道,并通过该通道将自身转运出去;这类蛋白质被称为自转运蛋白(autotransporters)。其他蛋白质则在一种独立的辅助蛋白(helper protein)的帮助下,通过V型通路分泌。
ABC蛋白质分泌通路(因其含有ATP结合盒(ATP binding cassette, ABC)而得名)在原核生物中普遍存在,即在革兰氏阳性菌、革兰氏阴性菌和古菌中均有发现。它有时也被称为I型蛋白质分泌通路(图4)。
在致病性革兰氏阴性菌中,它参与分泌毒素(如α-溶血素(α-hemolysin))、蛋白酶、脂肪酶和特定肽类。分泌的蛋白质通常含有C端分泌信号(C-terminal secretion signals),可引导新合成的蛋白质到达I型分泌机器。该机器贯穿质膜、周质空间和外膜。这些系统能一步将蛋白质跨过两层膜转运出去,从而绕过Sec依赖通路。
革兰氏阳性菌则使用一种改良版的I型系统来将蛋白质转运穿过质膜。对枯草芽孢杆菌(Bacillus subtilis)基因组的分析发现了77种ABC转运蛋白。这可能反映了ABC转运蛋白除了转运蛋白质外,还能转运多种溶质,包括糖和氨基酸,并能将药物从细胞内泵出。
几种革兰氏阴性病原菌拥有III型蛋白质分泌通路,这是另一种绕过Sec依赖通路的分泌系统。
大多数III型系统能将其攻击的动植物宿主细胞直接注入毒力因子(virulence factors)。这些毒力因子包括毒素、吞噬作用抑制剂、宿主细胞骨架重组刺激物以及宿主细胞凋亡(apoptosis)促进剂。
但在某些情况下,毒力因子只是被简单地分泌到胞外环境中。III型系统还能转运其他蛋白质,包括:①构成系统自身的部分蛋白质、②调控分泌过程的蛋白质、③协助分泌蛋白插入靶细胞的蛋白质。
III型系统结构复杂,常呈注射器状(图4)。其细长的针状部分从细胞表面伸出;圆柱形的基座连接着外膜和质膜,外观与鞭毛基体(flagellar basal body)有些相似。人们认为蛋白质可能通过一个易位通道(translocation channel)移动。
拥有III型系统的细菌的重要例子包括沙门氏菌(Salmonella)、耶尔森菌(Yersinia)、志贺氏菌(Shigella)、大肠杆菌(E. coli)、博德特氏菌(Bordetella)、铜绿假单胞菌(Pseudomonas aeruginosa)和欧文氏菌(Erwinia)。
IV型蛋白质分泌通路的独特之处在于,它既可用于分泌蛋白质,也可在细菌接合(conjugation)过程中将DNA从供体菌转移到受体菌。IV型系统由多种不同的蛋白质组成,与III型系统一样,这些蛋白质也形成注射器状结构。
参考文献
《Microbiology (Seventh Edition)》 | 微生物学,第七版
相关产品
HZB231810:大肠杆菌/Escherichia coli
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-05-11
编制人:叶凡
审稿人:小藻