从基础开始学习原核细胞04:细菌细胞壁
来源:武汉市灰藻生物科技有限公司 浏览量:772 发布时间:2026-05-09 22:10:41
1、引言
细胞壁位于质膜外侧,通常较为坚硬,它可以维持细胞形状、防止渗透裂解、抵御有毒物质,在病原菌中,还能增强致病性。
绝大多数原核生物都有细胞壁。少数无细胞壁的种类,也具备其他可替代其功能的结构。多种抗生素的作用靶点正是细胞壁,因此理解其结构至关重要。
2、细菌细胞壁结构概述
1884年,克里斯蒂安·革兰(Christian Gram)发明了革兰氏染色法。人们很快发现,多数细菌可根据染色反应分为两大类:革兰氏阳性菌呈紫色,革兰氏阴性菌呈粉红或红色。
直到透射电子显微镜出现,两类细菌的真实结构差异才得以明确。如图1所示,革兰氏阳性菌的细胞壁由一层均质的肽聚糖(peptidoglycan,又称胞壁质murein)构成,厚度为20至80纳米,位于质膜外侧。
相比之下,革兰氏阴性菌的细胞壁结构更复杂。其肽聚糖层仅厚2至7纳米,外覆一层7至8纳米厚的外膜(outer membrane)。
由于肽聚糖层更厚,革兰氏阳性菌对渗透压的抵抗力强于革兰氏阴性菌。微生物学家常将质膜及其外的所有结构统称为细胞包膜(cell envelope),包括质膜、细胞壁,以及荚膜等。
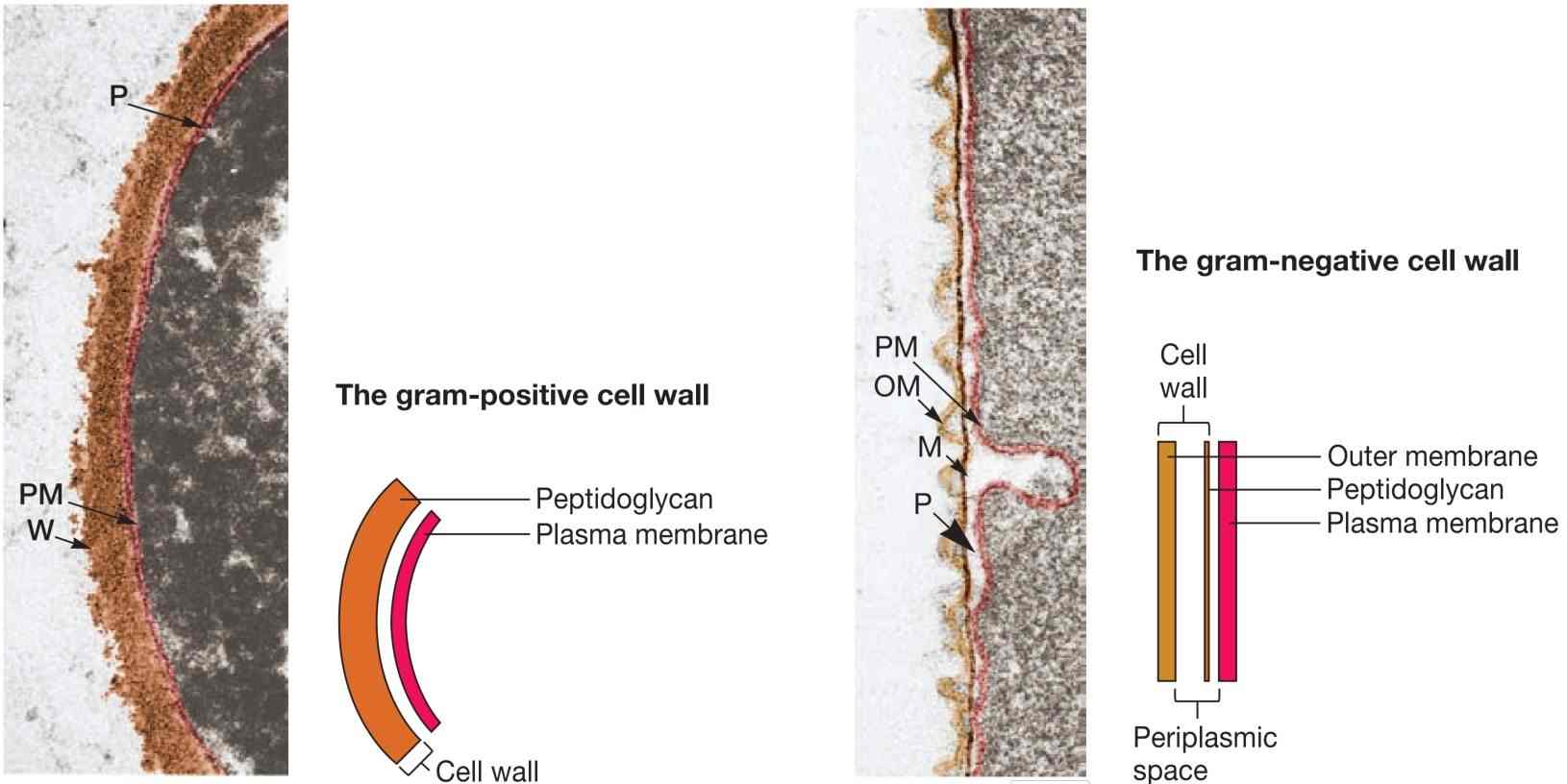
图1 革兰氏阳性与革兰氏阴性细胞壁。左侧为地衣芽孢杆菌(Bacillus licheniformis)的革兰氏阳性包膜,右侧为蛇形水螺菌(Aquaspirillum serpens)的革兰氏阴性电镜图。M:肽聚糖或胞壁质层;OM:外膜;PM:质膜;P:周质空间;W:革兰氏阳性肽聚糖壁。
在革兰氏阴性菌的电镜照片中,常可见质膜与外膜之间存在一个间隙,在革兰氏阳性菌中有时也能观察到质膜与细胞壁之间的类似间隙。此间隙称为周质空间(periplasmic space),其中的物质称为周质(periplasm)。
3、肽聚糖结构
肽聚糖(peptidoglycan)或胞壁质(murein)是一种巨大的网状聚合物,由许多相同的亚基组成。该聚合物包含两种糖衍生物:N-乙酰葡糖胺(N-acetylglucosamine, NAG)和N-乙酰胞壁酸(N-acetylmuramic acid, NAM,即N-乙酰葡糖胺的乳酸醚),以及多种氨基酸。
细菌肽聚糖中有三种氨基酸在一般细胞的结构蛋白中不常见:D-谷氨酸(D-glutamic acid)、D-丙氨酸(D-alanine)和内消旋二氨基庚二酸(meso-diaminopimelic acid)。D-型氨基酸的存在可防止细菌被大多数蛋白酶降解,因为这些酶通常只识别L-型氨基酸残基。
图2展示了多数革兰氏阴性菌及许多革兰氏阳性菌的肽聚糖亚基结构。该聚合物的主链由交替的N-乙酰葡糖胺(NAG)和N-乙酰胞壁酸(NAM)残基构成。一条由四个交替的D-型和L-型氨基酸组成的肽链,连接在N-乙酰胞壁酸的羧基上。许多细菌会用另一种二氨基氨基酸(通常是L-赖氨酸/L-lysine)替代内消旋二氨基庚二酸(见图3)。
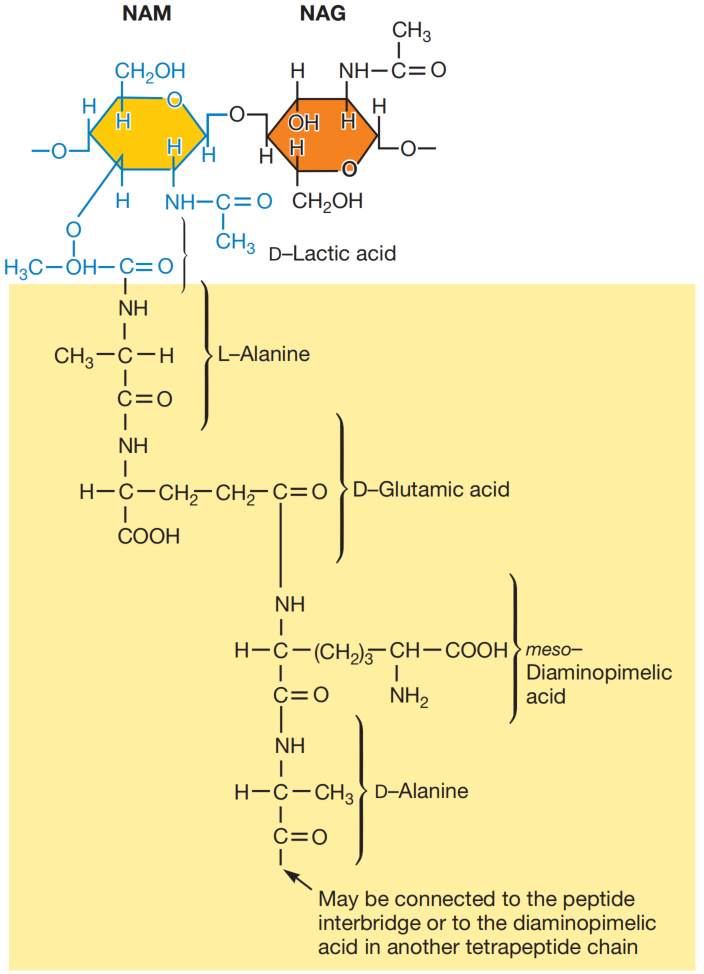
图2 肽聚糖亚基组成。大肠杆菌(E. coli)、多数其他革兰氏阴性菌及许多革兰氏阳性菌的肽聚糖亚基。NAG为N-乙酰葡糖胺。NAM为N-乙酰胞壁酸(NAG通过醚键连接乳酸)。四肽侧链由交替的D-型和L-型氨基酸组成,因内消旋二氨基庚二酸通过其L-碳原子连接。图中NAM及其连接的四肽链以不同色阶区分,以便清晰展示。
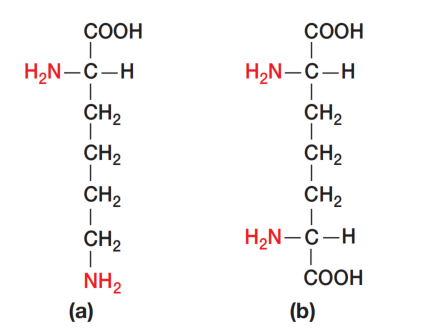
图3 肽聚糖中的二氨基氨基酸。(a) L-赖氨酸(L-Lysine)。(b) 内消旋二氨基庚二酸(meso-Diaminopimelic acid)。
为形成坚固的网状聚合物,相连的肽聚糖亚基链必须通过肽链间的交联(cross-links)连接起来。通常,末端D-丙氨酸的羧基直接与二氨基庚二酸的氨基相连,但有时也会通过一个肽间桥(peptide interbridge)连接(见图4)。
多数革兰氏阴性菌的细胞壁肽聚糖不含肽间桥。这种交联最终形成一个巨大的肽聚糖囊(sac),实际上是一个致密且相互连接的网络(见图5)。从革兰氏阳性菌中分离出的这类囊体,足以保持其形状和完整性(见图6)。同时,它们也具有多孔性、弹性和一定的延展性。
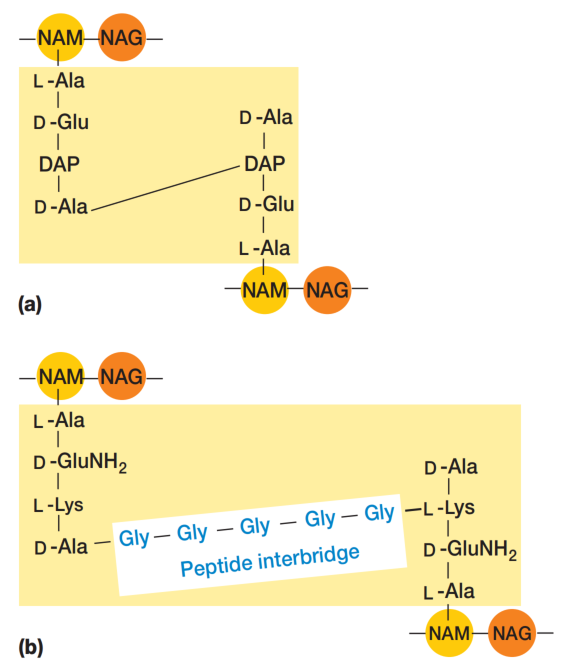
图4 肽聚糖交联。(a) 大肠杆菌(E. coli)肽聚糖,具有直接交联,是许多革兰氏阴性菌的典型结构。(b) 金黄色葡萄球菌(Staphylococcus aureus)肽聚糖S. aureus为革兰氏阳性菌。NAM为N-乙酰胞壁酸。NAG为N-乙酰葡糖胺。Gly为甘氨酸(glycine)。为清晰起见,图中多糖链被画成相对排列,但实际上两条并排的链即可相互连接(见图5)。
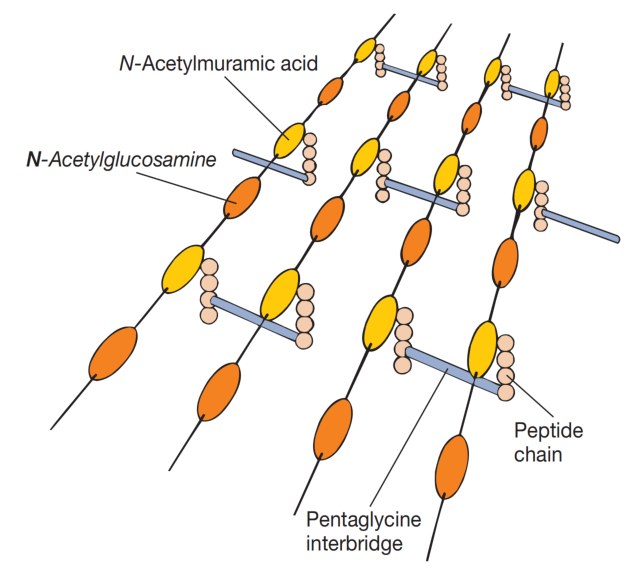
图5 肽聚糖结构。一种肽聚糖模型的示意图。图中展示了多糖链、四肽侧链和肽间桥。
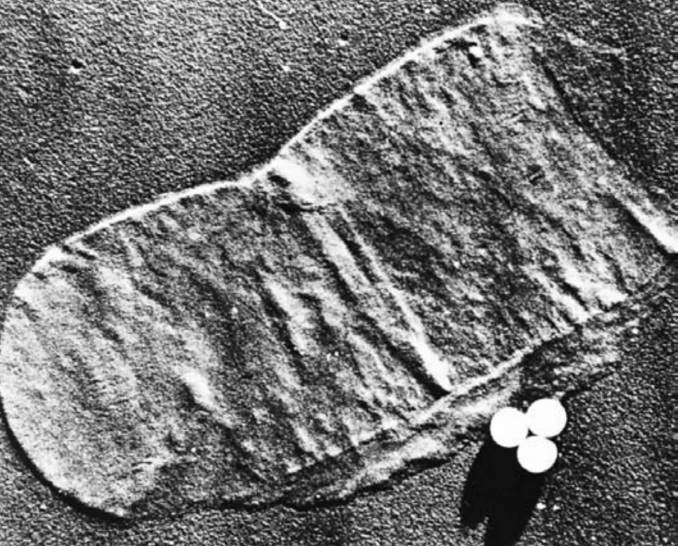
图6 分离的革兰氏阳性细胞壁。来自革兰氏阳性菌巨大芽孢杆菌(Bacillus megaterium)的肽聚糖壁。乳胶微球的直径为0.25微米。
4、革兰氏阳性细胞壁
革兰氏阳性菌的细胞壁通常很厚,主要由肽聚糖构成。其肽聚糖常含有肽间桥(见图5和图7)。
此外,革兰氏阳性细胞壁通常含有大量磷壁酸(teichoic acids)。磷壁酸是由甘油(glycerol)或核糖醇(ribitol)通过磷酸基团连接而成的聚合物(见图7和图8)。D-丙氨酸(D-alanine)等氨基酸或葡萄糖(glucose)等糖类会连接到甘油和核糖醇基团上。
磷壁酸通过共价键连接到肽聚糖本身或质膜脂质上。后者被称为脂磷壁酸(lipoteichoic acids)。磷壁酸似乎延伸至肽聚糖表面,因其带负电荷,有助于赋予革兰氏阳性细胞壁负电性。这些分子的功能尚不明确,但它们可能对维持细胞壁结构很重要。革兰氏阴性菌不含磷壁酸。
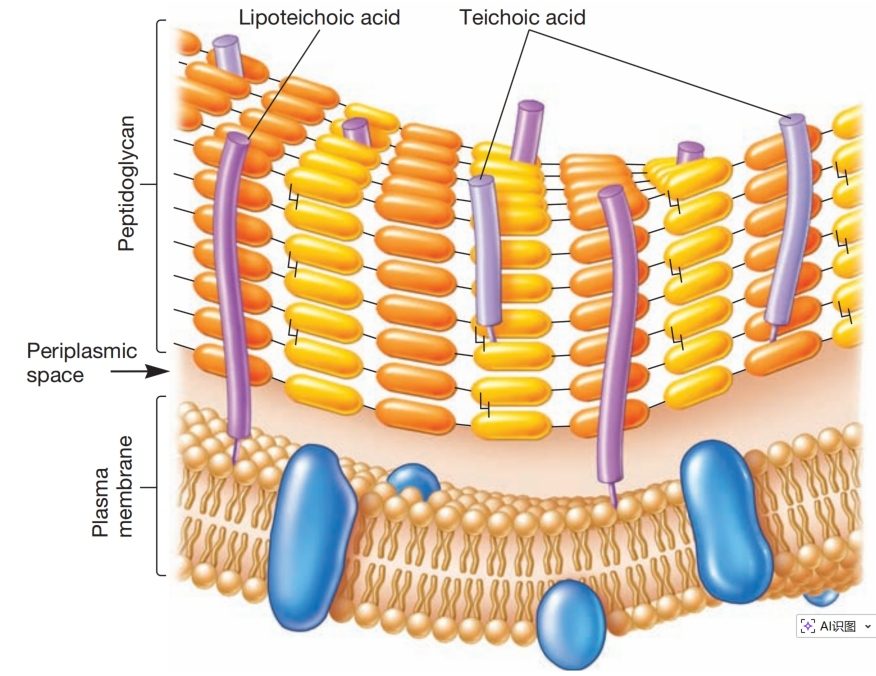
图7 革兰氏阳性包膜。
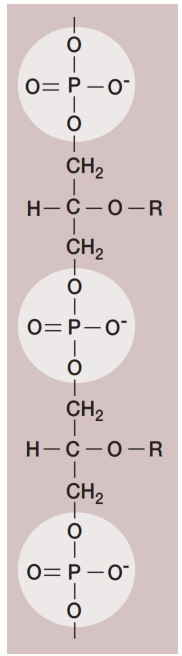
图8 磷壁酸结构。一段由磷酸、甘油和侧链R组成的磷壁酸。R可代表D-丙氨酸、葡萄糖或其他分子。
革兰氏阳性菌的周质空间(若存在)位于质膜与细胞壁之间,比革兰氏阴性菌的小。即使革兰氏阳性菌没有明显的周质空间,也可能存在周质。
周质中的蛋白质相对较少。这可能是因为肽聚糖囊是多孔的,细胞分泌的任何蛋白质通常都能穿过它。革兰氏阳性菌分泌的酶称为胞外酶(exoenzymes),常用于降解大分子营养物质,使其能被转运过质膜。留在周质空间的蛋白质通常附着在质膜上。
葡萄球菌(Staphylococci)和大多数革兰氏阳性菌在其细胞壁肽聚糖表面有一层蛋白质。这些蛋白质参与细胞与环境的相互作用。
有些蛋白质通过非共价方式结合到肽聚糖、磷壁酸或其他受体上。例如,S层蛋白(S-layer proteins),通过非共价方式结合到分散在细胞壁中的聚合物上。参与肽聚糖合成与周转的酶似乎也以非共价方式与细胞壁相互作用。
其他表面蛋白则通过共价键连接到肽聚糖上。许多共价连接的蛋白质(如致病性链球菌的M蛋白)具有毒力作用,例如帮助粘附宿主组织或干扰宿主防御。在葡萄球菌中,这些表面蛋白通过共价键连接到细胞壁肽聚糖的五甘氨酸桥(pentaglycine interbridge)上。
一种名为分选酶(sortase)的酶催化这些表面蛋白与革兰氏阳性肽聚糖的连接。分选酶附着在细菌细胞的质膜上。
5、革兰氏阴性细胞壁
从图1可以看出,革兰氏阴性细胞壁比革兰氏阳性壁复杂得多。紧邻质膜的薄肽聚糖层,两侧被周质空间包围,其重量可能不超过细胞壁总重的5%至10%。在大肠杆菌(E. coli)中,肽聚糖层厚度约为2纳米,仅含一到两层肽聚糖。
革兰氏阴性菌的周质空间也与革兰氏阳性菌显著不同。其大小范围从1纳米到71纳米不等。一些新研究表明,它可能占细胞总体积的20%至40%,通常宽30至70纳米。
当细胞壁被破坏或移除而不影响下方质膜时,周质酶和其他蛋白质会被释放出来,便于研究。
一些周质蛋白参与营养获取,例如水解酶和转运蛋白。
一些周质蛋白参与能量守恒。例如,将硝酸盐转化为氮气的反硝化细菌,以及利用无机分子作为能源的化能无机自养菌(chemolithotrophs),其周质中含有电子传递蛋白。
其他周质蛋白参与肽聚糖合成及有毒化合物的修饰,以保护细胞免受伤害。
外膜位于薄肽聚糖层之外(见图9和图10),并通过两种方式与细胞连接。
第一种是通过Braun脂蛋白(Braun's lipoprotein),这是外膜中最丰富的蛋白质。这种小脂蛋白通过共价键连接到下方的肽聚糖,并通过其疏水端嵌入外膜。外膜和肽聚糖通过这种脂蛋白牢固连接,可以作为一个整体被分离出来。
第二种连接机制涉及外膜和质膜之间的多个粘附位点。在这两个位点,两种膜似乎直接接触。在大肠杆菌中,可以看到两个膜之间有20至100纳米的接触区域。粘附位点可能是直接接触区,也可能是真正的膜融合区。有人提出,物质可以直接通过这些粘附位点进入细胞,而无需穿过周质。
外膜最不寻常的成分可能是其脂多糖(lipopolysaccharides, LPSs)。这些大而复杂的分子同时含有脂质和碳水化合物,由三部分组成:(1) 脂质A(lipid A),(2) 核心多糖(core polysaccharide),(3) O侧链(O side chain)。
沙门氏菌(Salmonella)的LPS被研究得最多,其一般结构如下所述(见图11)。脂质A区域包含两个氨基葡萄糖(glucosamine)糖衍生物,每个都连接有三个脂肪酸和一个磷酸基或焦磷酸基。
脂肪酸将脂质A锚定在外膜上,而LPS分子的其余部分则从表面伸出。核心多糖连接在脂质A上。在沙门氏菌中,它由10个糖组成,其中许多结构特殊。
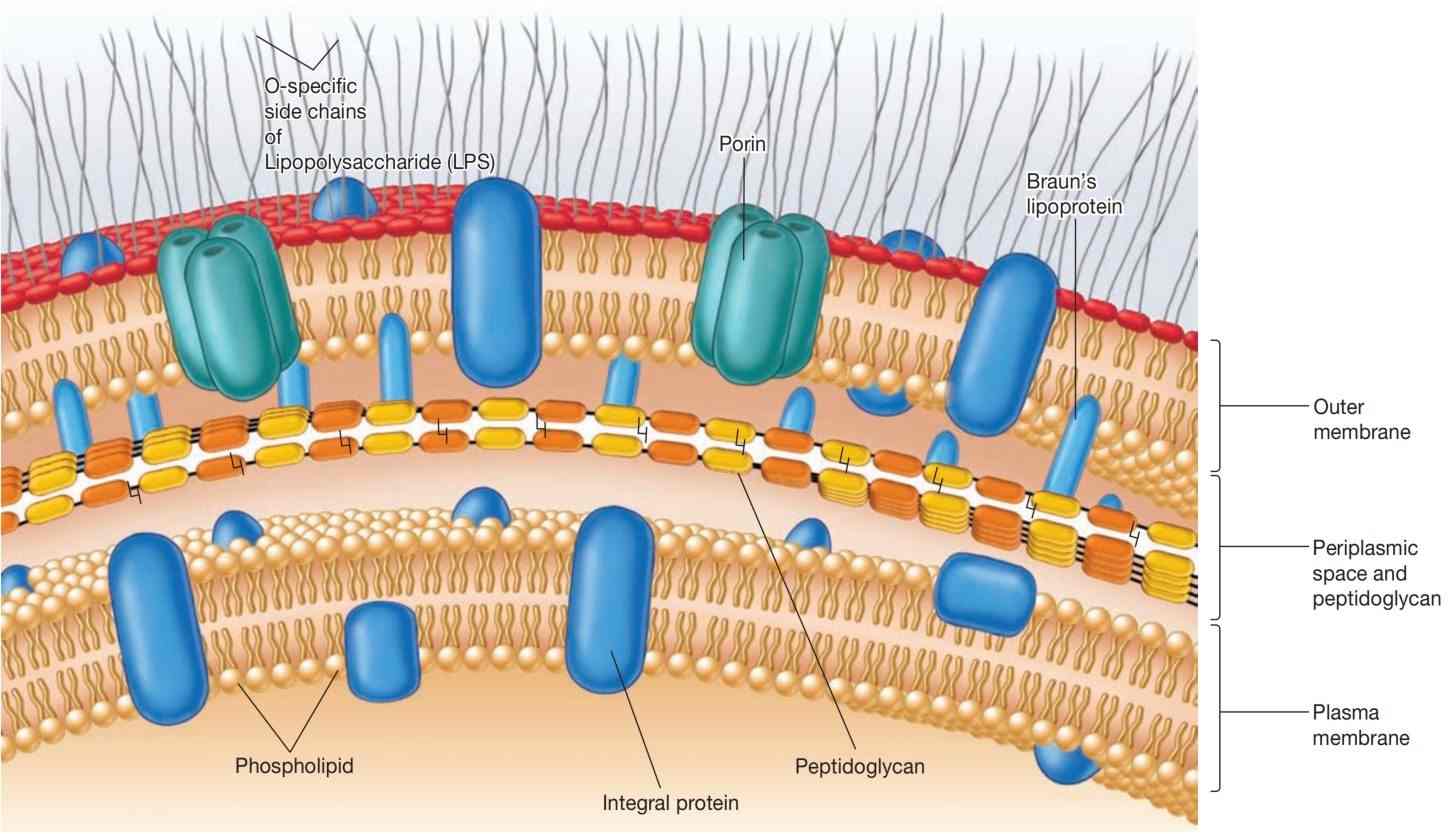
图9 革兰氏阴性包膜。
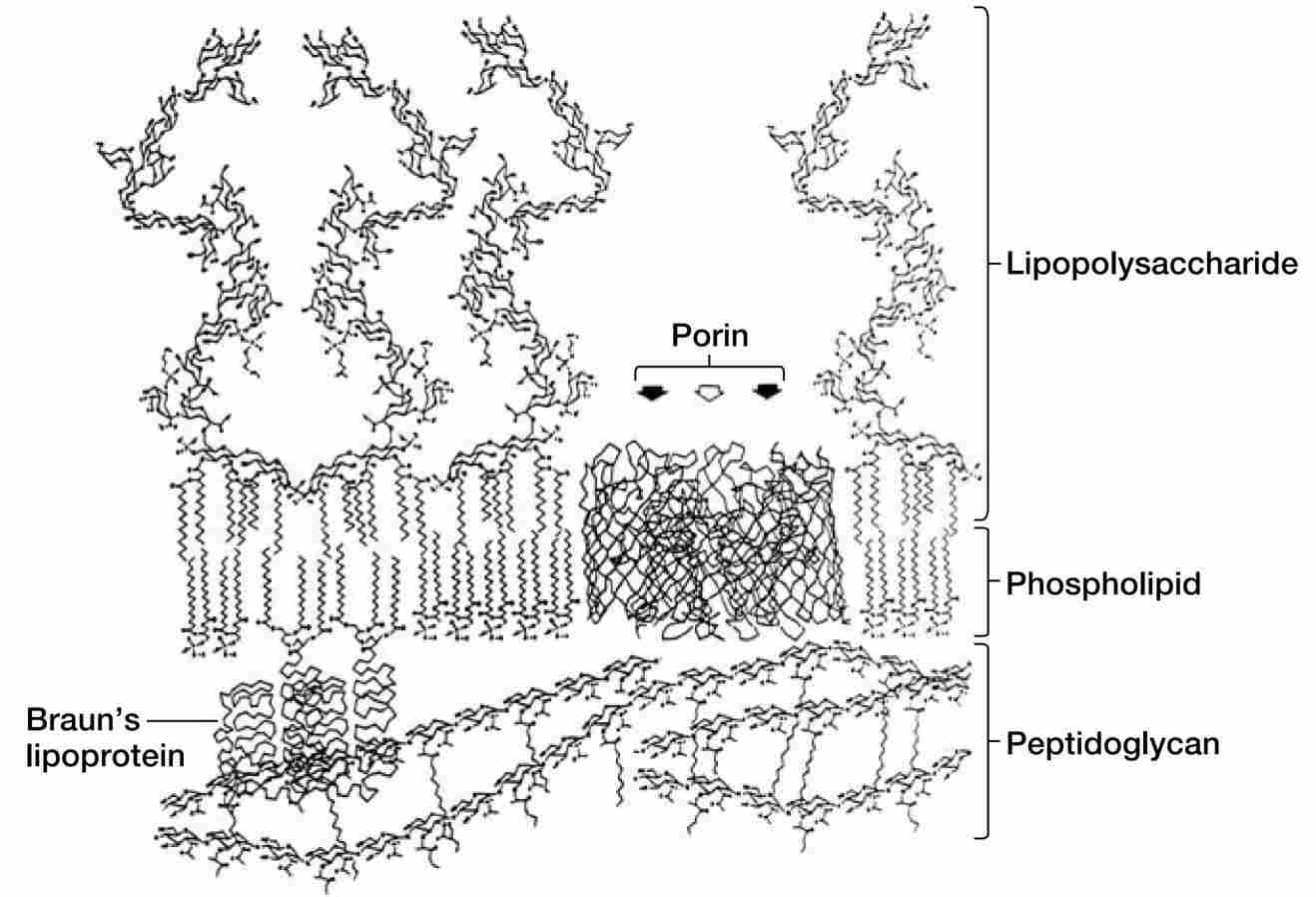
图10 大肠杆菌外膜及相关结构的化学模型。此横截面为比例图。孔蛋白OmpF在三聚体蛋白复合物的前方有两个通道(实心箭头),后方有一个通道(空心箭头)。LPS分子可以比图中所示的更长。
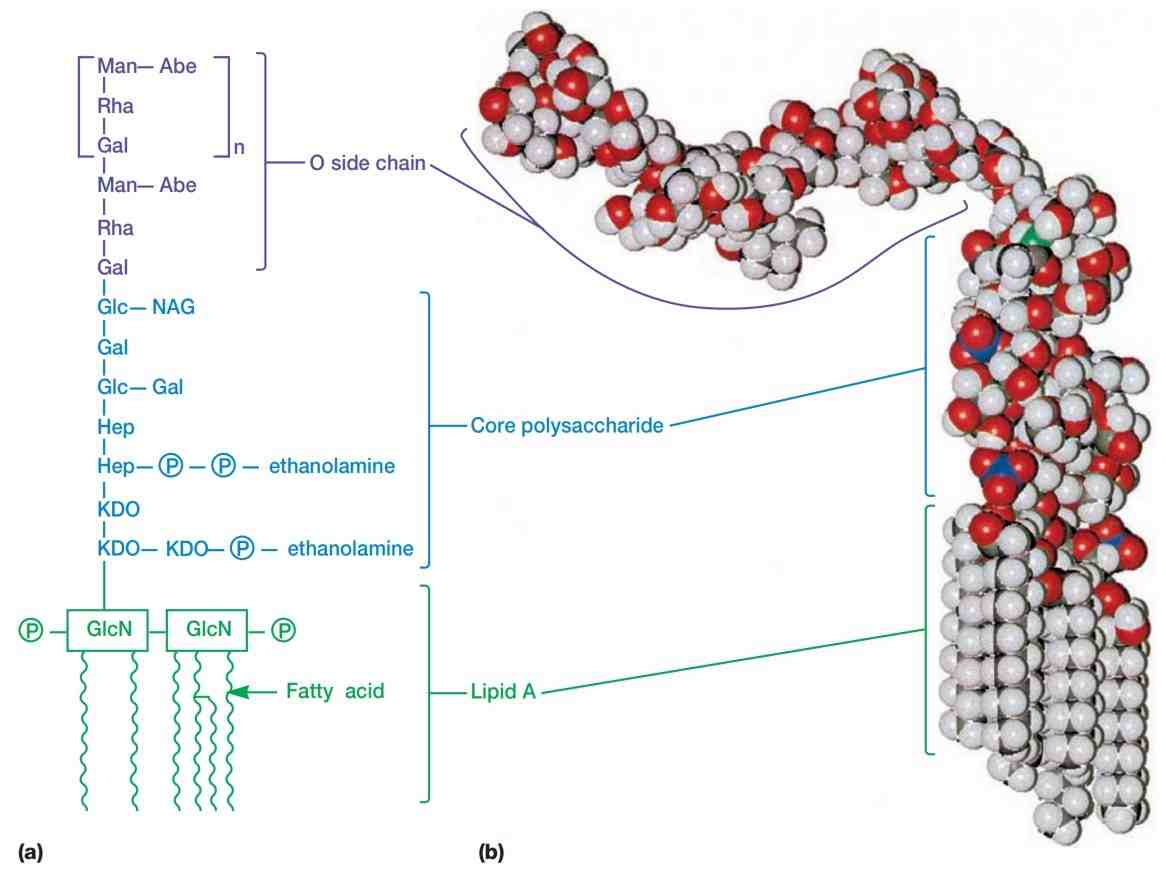
图11 脂多糖结构。(a) 沙门氏菌的脂多糖。LPS。此略简化的图示展示了一种LPS形式。缩写:Abe,阿比糖(abequose);Gal,半乳糖(galactose);Glc,葡萄糖(glucose);GlcN,氨基葡萄糖(glucosamine);Hep,庚酮糖(heptulose);KDO,2-酮-3-脱氧辛酸(2-keto-3-deoxyoctonate);Man,甘露糖(mannose);NAG,N-乙酰葡糖胺(N-acetylglucosamine);P,磷酸(phosphate);Rha,L-鼠李糖(L-rhamnose)。脂质A埋在外膜中。(b) 大肠杆菌脂多糖的分子模型。脂质A和核心多糖是直的;在此模型中,O侧链呈一定角度弯曲。
LPS有许多重要功能。由于核心多糖通常含有带电糖和磷酸基(见图11),LPS有助于赋予细菌表面负电荷。作为外膜外小叶的主要成分,脂质A也有助于稳定外膜结构。LPS可能有助于细菌附着在表面并形成生物膜。
LPS的一个主要功能是帮助形成通透性屏障。LPS的几何结构(见图11b)及相邻LPS分子间的相互作用被认为能限制胆汁盐、抗生素及其他有毒物质的进入,从而保护细菌。
LPS还在保护致病性革兰氏阴性菌免受宿主防御方面发挥作用。LPS的O侧链也被称为O抗原,因为它能引发免疫反应。这种反应涉及产生抗体,这些抗体会结合引发反应的特定LPS菌株。
然而,许多革兰氏阴性菌能够快速改变其O侧链的抗原性质,从而逃避宿主防御。重要的是,LPS的脂质A部分通常具有毒性;因此,LPS可作为内毒素(endotoxin),引起革兰氏阴性菌感染的部分症状。如果细菌进入血液,LPS内毒素可导致败血性休克,目前尚无直接治疗方法。
尽管LPS在形成通透性屏障方面起作用,但外膜比质膜更具通透性,允许葡萄糖等小分子单糖通过。这是由于孔蛋白(porin proteins)的存在(见图9和图10)。
大多数孔蛋白在外膜中聚集形成三聚体(见图9和图12)。每个孔蛋白贯穿外膜,形状大致呈管状;其狭窄的通道允许分子量小于约600至700道尔顿的分子通过。
然而,像维生素B12这样的大分子也能穿过外膜。这些大分子不通过孔蛋白,而是由特定的载体将其转运过外膜。
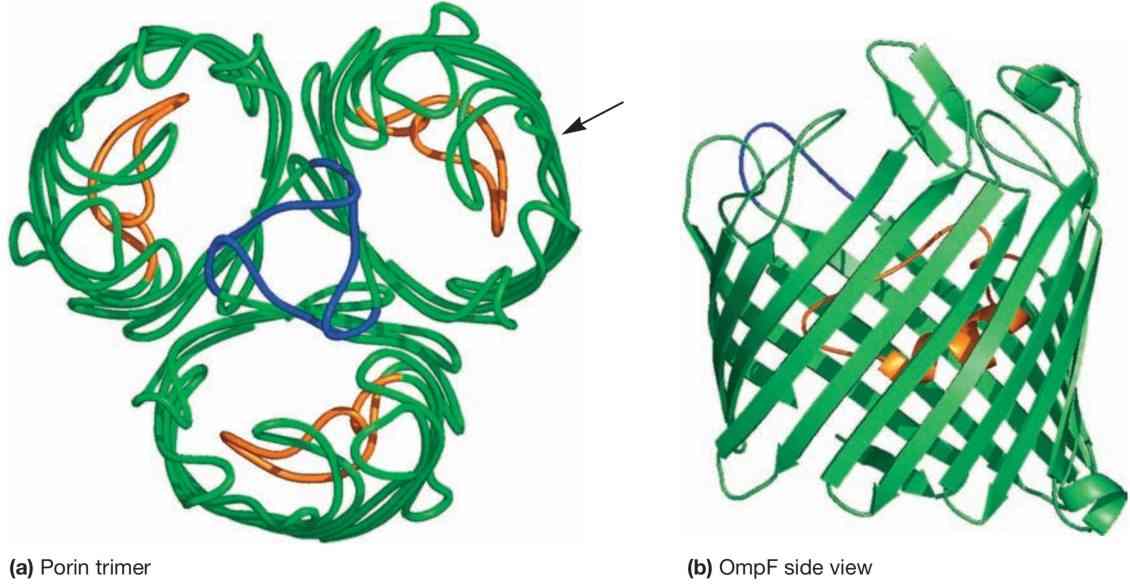
图12 孔蛋白(Porin Proteins)。大肠杆菌(E. coli)OmpF孔蛋白的两种视图。(a) 从外膜外表面俯视的孔蛋白三聚体结构(即顶视图)。构成该蛋白复合物的三个孔蛋白各自形成一个通道。每个孔蛋白可分为三个环:绿色环形成通道,蓝色环与其他孔蛋白相互作用以帮助形成三聚体,橙色环使通道变窄。箭头指示了(b)图中从侧面观察的孔蛋白分子区域。(b) 孔蛋白单体的侧视图,展示了孔蛋白特有的β-桶(β-barrel)结构。
6、革兰氏染色的机理
革兰氏染色反应结果有多种解释。目前认为,革兰氏阳性菌与阴性菌的差异源于其细胞壁的物理特性。
如果去除革兰氏阳性菌的细胞壁,它们会染成革兰氏阴性。同样,天生无细胞壁的支原体(mycoplasmas)也染成革兰氏阴性。肽聚糖本身不被染色,而是作为通透性屏障,防止结晶紫(crystal violet)流失。
染色过程中,细菌首先用结晶紫染色,再用碘液处理以促进染料滞留。随后用乙醇处理时,酒精会使革兰氏阳性菌厚实的肽聚糖层孔隙收缩。因此,在短暂的脱色步骤中,染料-碘复合物得以保留,细菌保持紫色。
相比之下,革兰氏阴性菌的肽聚糖层很薄,交联度较低,孔隙更大。酒精处理还可能从其外膜中提取足够多的脂质,进一步增加通透性。因此,酒精更容易将紫色的结晶紫-碘复合物从革兰氏阴性菌中洗脱。这样,它们就能被复染剂沙黄(safranin)轻易地染成红色或粉色。
7、细胞壁与渗透保护
微生物有多种机制应对渗透压变化。当细胞内外溶质浓度不同时,就会产生渗透压。适应性反应旨在平衡溶质浓度。
但在某些情况下,渗透压会超过细胞的适应能力。此时,细胞壁提供额外保护。当细胞处于低渗溶液(溶质浓度低于细胞质)中时,水会进入细胞,导致其膨胀。
没有细胞壁,质膜承受的压力会过大,最终破裂,细胞裂解(lysis)。相反,在高渗溶液中,水会流出,细胞质会皱缩,这个过程称为质壁分离(plasmolysis)。
细胞壁的保护作用在用溶菌酶(lysozyme)或青霉素(penicillin)处理细菌时表现得最为明显。
溶菌酶通过水解连接N-乙酰胞壁酸(N-acetylmuramic acid)和N-乙酰葡糖胺(N-acetylglucosamine)的键来攻击肽聚糖(见图2)。青霉素则通过不同机制起作用,它抑制肽聚糖的合成。
如果细菌在低渗溶液中接受这两种物质之一的处理,它们会裂解。但如果在等渗溶液中,它们可以存活并正常生长。
对于革兰氏阳性菌,经溶菌酶或青霉素处理后,细胞壁会完全丧失,细胞变成原生质体(protoplast)。当革兰氏阴性菌暴露于溶菌酶或青霉素时,肽聚糖层会丢失,但外膜仍然存在。这些细胞被称为球状体(spheroplasts)。
由于缺乏完整的细胞壁,原生质体和球状体对渗透压敏感。如果将它们转移到稀释溶液中,会因水分不受控制地涌入而裂解(见图13)。
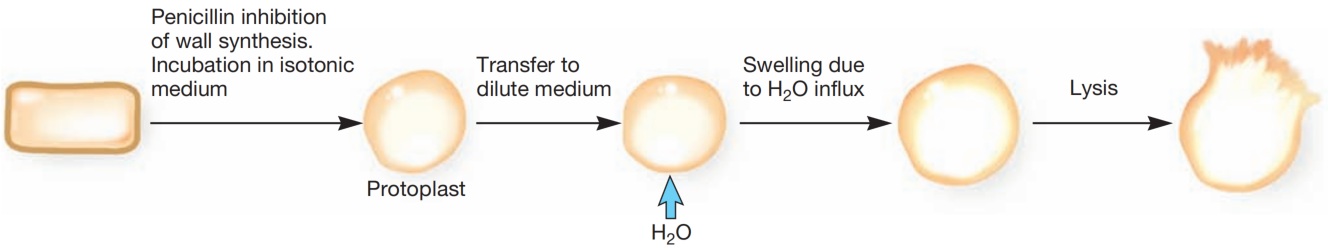
图13 原生质体的形成与裂解。在等渗培养基中用青霉素孵育诱导原生质体形成。转移到稀释培养基会导致裂解。
尽管大多数细菌需要完整的细胞壁才能生存,但有些细菌完全没有细胞壁。例如,支原体(mycoplasmas)缺乏细胞壁,对渗透压敏感,却常能在稀释培养基或陆地环境中生长。这是因为它们的质膜比有壁细菌的质膜更能抵抗渗透压。确切原因尚不清楚,但许多支原体膜中存在甾醇(sterols),这可能提供了额外的强度。由于没有坚硬的细胞壁,支原体往往呈多形性(pleomorphic)或形状多变。
参考文献
《Microbiology (Seventh Edition)》 | 微生物学,第七版
相关产品
HZB231810:大肠杆菌/Escherichia coli
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-05-09
编制人:叶凡
审稿人:小藻