微生物降解纤维素:从酶系协同到分子机制的深度解析
来源:武汉市灰藻生物科技有限公司 浏览量:449 发布时间:2026-03-30 16:16:41
引言
纤维素作为生物圈中,储量最丰富的有机聚合物,不仅是植物细胞壁的骨架支撑,更是全球碳循环中至关重要的碳库。
然而,其独特的β-1,4-糖苷键连接方式,及高度结晶的超分子结构,赋予了纤维素极强的化学惰性与物理稳定性,使其成为自然界中难以被直接利用的“顽固”底物。
尽管人类及多数动物缺乏代谢纤维素的能力,但微生物界却演化出了一套精妙而高效的降解系统。
从土壤中的腐生真菌到反刍动物瘤胃内的厌氧细菌,各类纤维素降解微生物通过分泌复杂的酶系,将难溶的纤维素转化为可发酵的单糖,从而驱动着生态系统的能量流动。
近年来,随着生物能源与合成生物学的兴起,深入解析这一生物转化过程——包括纤维素的精细结构特征、产酶微生物的多样性、以及水解与氧化并存的分子机制——已成为突破生物质转化瓶颈的关键。
本文将系统梳理微生物降解纤维素的全过程,从基础的结构认知出发,深入探讨酶系的协同作用及其背后的生化机理,为理解自然界的碳循环及开发新型生物炼制技术提供理论基石。
什么是纤维素?
纤维素是一种有机多糖,由数百个通过β-糖苷键连接的D-葡萄糖单元构成的线性长链聚合物。
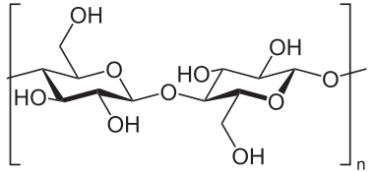
图1、纤维素是由D-葡萄糖单元通过β(1→4)-糖苷键连接形成的线性聚合物(图片来源:NEUROtiker)
作为生物圈中含量最丰富的胞外结构多糖及有机聚合物,纤维素广泛存在于所有陆生植物中,但在肉类、蛋类、鱼类及乳制品中完全缺失。尽管人体无法代谢纤维素,但其作为膳食纤维的重要来源,对健康至关重要。
在植物界,纤维素是分布最广的碳水化合物,约占植被总碳量的50%。它主要存在于植物细胞壁中,是维持植物体结构完整性的关键组分。
所有能合成纤维素的生物(包括细菌、藻类、被囊动物及高等植物)均含有纤维素合酶蛋白,该酶催化葡聚糖链的聚合反应。
在自然界中,纤维素是细菌、真菌、植物、原生生物以及多种无脊椎动物(如昆虫、甲壳类、环节动物、软体动物和线虫)的重要食物来源。
纤维素独特的半结晶聚集态使其在水环境中仍能保持结构稳定,赋予了植物细胞卓越的机械强度。
作为一种葡萄糖衍生物的均聚物,纤维素是优质的可发酵糖来源。目前,纤维素已被作为能源作物广泛种植,用于生产乙醇、醚类、乙酸等化工产品。
得益于高等植物持续的光合作用循环,全球每年合成的纤维素总量高达约10亿吨。
纤维素呈纤维状、刚性白色固体,不溶于水,但可溶于氨性氢氧化铜溶液。尽管不溶于水,纤维素能吸收水分并增加粪便体积,从而促进其排出体外。
纤维素的精细结构
纤维素分子量介于20万至200万道尔顿之间,对应每个分子包含1,250至12,500个葡萄糖残基。其分子一端为含C4-OH基团的非还原端(D-葡萄糖单元),另一端为含C1-OH基团的还原端。
图2、微生物降解纤维素过程,包含酶系、步骤与机制(制图工具:BioRender.com)
纤维素链的形成机制为:一个β-D-葡萄糖分子C1位的糖苷羟基与相邻β-D-葡萄糖分子C4位的醇羟基脱去一分子水,形成β-1,4-糖苷键。纤维二糖是纤维素的基本重复单元。
纤维素的整体结构源于相邻纤维素链及片层间通过氢键和范德华力结合,形成平行排列。这种排列方式造就了纤维素的结晶结构,使其具有笔直、稳定的超分子纤维形态,表现出极高的抗张强度和较低的可及性。
除葡萄糖单元通过β-1,4-糖苷键连接外,纤维素结构与直链淀粉相似。这种β-1,4-糖苷键构型使得线性葡聚糖链中每隔一个葡萄糖残基相对于邻位旋转180°。
纤维素分子具有极高的稳定性,在25°C条件下,其β-糖苷键断裂的半衰期长达500万至800万年。根据结构及可及性差异,纤维素可分为结晶型与非结晶型、可及型与不可及型。
木材中的纤维素大部分为高度结晶态(结晶区占比约65%),其余部分因堆积密度较低而呈现无定形或非结晶结构。
"纤维素可及性"用于定义水和微生物对纤维素的利用程度。结晶纤维素表面通常具有较高的可及性,而其内部结构则较难被接触。
什么是纤维素酶?
纤维素酶是一类催化纤维素降解为寡糖、纤维二糖及葡萄糖的酶系总称。
这类酶主要由真菌和细菌产生,在纤维素水解过程中发挥关键作用。作为自然界和工业领域的重要酶类,纤维素酶通过降解难溶性纤维素为可溶性形式,积极参与全球碳循环。
尽管纤维素酶家族内存在七种不同的蛋白质折叠类型,但它们均能特异性水解单一底物——纤维素,展现出结构的多样性与功能的专一性。
完整的纤维素酶系统包含三种核心酶:外切β-1,4-葡聚糖酶、内切β-1,4-葡聚糖酶及β-1,4-葡萄糖苷酶。这些酶通过协同作用依次催化,将纤维素逐步降解为可利用的能源物质——葡萄糖。
根据作用模式,纤维素酶通常分为四大类:外切葡聚糖酶、内切葡聚糖酶、β-葡萄糖苷酶及纤维二糖水解酶。尽管各类酶在结构和作用机制上存在差异,但在特定条件下可 sequential 协作以生成目标产物。
此外,不同生物来源的纤维素酶(如真菌源与细菌源)在结构与功能上显著不同。例如,真菌纤维素酶通常在C端含有碳水化合物结合模块(CBM),通过短肽连接区与N端的催化结构域相连,而细菌纤维素酶则不具备此典型结构特征。
纤维素降解微生物概览
多年来,研究人员已鉴定出种类繁多的纤维素降解微生物,主要包括真菌和细菌。不同微生物产生的纤维素酶在结构与作用模式上存在显著差异。纤维素降解真菌
真菌是有机物质(尤其是纤维素底物)分解过程中最活跃的参与者之一。产纤维素酶的真菌分布广泛,涵盖子囊菌门(如里氏木霉 Trichoderma reesei)、担子菌门(如沼泽卧孔菌 Fomitopsis palustris)以及少数厌氧真菌物种。
在各类真菌中,软腐菌以产纤维素酶能力著称,其中木霉属(Trichoderma)的研究最为深入。其他知名的产纤维素酶软腐菌包括黑曲霉(Aspergillus niger)、尖孢镰刀菌(Fusarium oxysporum)和粗糙脉孢菌(Neurospora crassa)等。
除软腐菌外,褐腐菌和白腐菌也积极参与纤维素降解,但其作用机制截然不同:
• 褐腐菌:由于缺乏外切葡聚糖酶,主要在木材边材腐朽早期活跃水解纤维素。代表菌种包括Poria placenta、Lenzites trabea、Coniophora puteana及Tyromyces palustris。
• 白腐菌:主要参与木质纤维素的降解,代表菌种有黄孢原毛平革菌(Phanerochaete chrysosporium)、嗜热丝孢菌(Sporotrichum thermophile)和彩绒革盖菌(Trametes versicolor)。
在厌氧纤维素降解真菌中,研究较多的包括新美鞭菌(Neocallimastix frontalis)、普通派罗霉(Piromyces communis)及奥平霉属(Orpinomyces)物种。纤维素降解细菌
纤维素降解细菌通常产生的纤维素酶量较少,其降解过程往往依赖多酶复合体集群完成。目前报道的细菌纤维素酶主要来源于芽孢杆菌属(Bacillus)、不动杆菌属(Acinetobacter)、生孢噬纤维菌属(Cellulomonas)及梭菌属(Clostridium)。
约90%-95%的细菌纤维素酶活性由好氧细菌在有氧条件下产生,其余10%则由多样化的厌氧菌群在缺氧环境中完成。
此外,部分瘤胃细菌也能产生降解细胞壁组分的纤维素酶,代表性菌种包括产琥珀酸纤维杆菌(Fibrobacter succinogenes)、白色瘤胃球菌(Ruminococcus albus)、假单胞菌属(Pseudomonas)、变形杆菌属(Proteus)及葡萄球菌属(Staphylococcus)。某些嗜热细菌(如Anoxybacillus sp.、Geobacillus sp.及拟杆菌属)也表现出显著的纤维素酶活性。
纤维素降解关键酶系
参与纤维素降解的酶统称为纤维素酶,根据催化反应类型可分为五大类:
1. 内切葡聚糖酶(Endoglucanase)
内切葡聚糖酶属于内切纤维素酶,随机攻击纤维素分子非结晶区的内部键位,切断分子内的β-1,4-糖苷键。其主要功能是缩短纤维素链长度,为其他酶的作用创造位点。
2. 外切葡聚糖酶(Exoglucanases)
外切葡聚糖酶属于外切纤维素酶,特异性水解纤维素链的还原端或非还原端。其主要产物为纤维二糖,可进一步水解为单糖单元。该类酶作用于内切酶处理后形成的四糖和二糖片段,包括两类:
• 1,4-β-D-葡聚糖葡萄糖水解酶:从β-葡聚糖和细胞糊精中释放D-葡萄糖;
• 1,4-β-D-葡聚糖纤维二糖水解酶:以渐进方式从β-葡聚糖中释放D-纤维二糖。
3. 纤维二糖酶(Cellobiases)
纤维二糖酶作用于纤维二糖单元(包括二糖、三糖及四糖),将其转化为单糖。因其最终生成独立葡萄糖单元,亦被称为β-葡萄糖苷酶。
4. 氧化性纤维素酶(Oxidative cellulases)
此类酶通过自由基反应将纤维素解聚为小分子单元。例如,纤维二糖脱氢酶可催化多种形态物质转化为纤维二糖,便于后续被纤维二糖酶利用。
5. 纤维二糖磷酸化酶(Cellobiose phosphorylases)
与纤维二糖酶类似,但其水解聚合物的过程依赖磷酸盐而非水分子参与。
纤维素的有氧与厌氧降解机制
1. 有氧降解
有氧纤维素水解依赖三类酶的协同作用:内切葡聚糖酶(1,4-β-D-葡聚糖4-葡聚糖水解酶)、外切葡聚糖酶及β-葡萄糖苷酶(β-D-葡萄糖苷葡萄糖水解酶)。该过程逐步释放可溶性细胞糊精及各类糖苷中的D-葡萄糖单元。
有氧纤维素酶具有高浓度分泌特性,并按顺序发挥作用:
1. 内切酶首先攻击纤维素纤维的无定形区,形成作用位点;
2. 外切酶继而从纤维更结晶的区域水解出纤维二糖单元;
3. β-葡萄糖苷酶最终水解纤维二糖,生成单体葡萄糖。
2. 厌氧降解
厌氧细菌纤维素酶解聚结晶纤维素的机制尚不明确,但已知其与有氧水解存在本质区别。多数厌氧微生物的纤维素酶组装成大型多蛋白复合体——纤维素酶体(cellulosomes)。
该结构使细胞与底物紧密接触,最大限度减少水解产物的扩散损失。
影响纤维素降解的关键因素
土壤或其他环境中的纤维素降解速率受多种因素调控:1. 矿物质可利用性
营养物质与矿物质的供应直接影响纤维素酶及其他蛋白质的合成。养分增加可显著提升降解速率。2. 温度
纤维素降解可在0°C至65°C范围内进行(嗜冷与嗜热微生物均具此能力),但最适温度为中温区间(25-30°C)。3. 通气条件
氧气可用性同时影响水解速率与机制:
• 有氧条件:三类酶 sequential 协作完成葡萄糖转化;
• 厌氧条件:多酶复合体作用效率相对较低。4. pH值
酸性土壤中纤维素降解速率略高于碱性或中性土壤:
• 酸性环境:真菌主导分解过程;
• 中性至碱性环境:细菌与放线菌成为主要降解者。5. 有机质含量
有机质可作为底物促进降解,但若体系中仅含纤维素,水解速率反而下降。添加少量易分解有机质可刺激微生物生长,从而加速降解进程。6. 木质素影响
木质素与纤维素结构紧密关联,其存在会抑制纤维素降解速率。
纤维素降解的三大核心步骤
纤维素的生物降解通常遵循三个连续且协同的步骤:
1. 内切葡聚糖酶的水解作用
这是降解过程的起始阶段。内切葡聚糖酶随机攻击纤维素微纤丝的内部位点。该酶在聚合物链的非特定位置进行切割,将长链纤维素降解为较短的片段,从而显著降低纤维素链的长度并增加其末端数量。
2. 外切葡聚糖酶的水解作用
随后,外切葡聚糖酶作用于上述产生的较小片段。它们特异性地攻击片段末端的还原端或非还原端,将其进一步水解为四糖或二糖(主要是纤维二糖)单元。
3. β-葡萄糖苷酶的水解作用
这是降解的最后一步。β-葡萄糖苷酶(亦称纤维二糖酶)作用于纤维二糖等二糖单元,将其水解为单体葡萄糖分子。至此,纤维素被完全转化为可被微生物直接利用的游离葡萄糖。
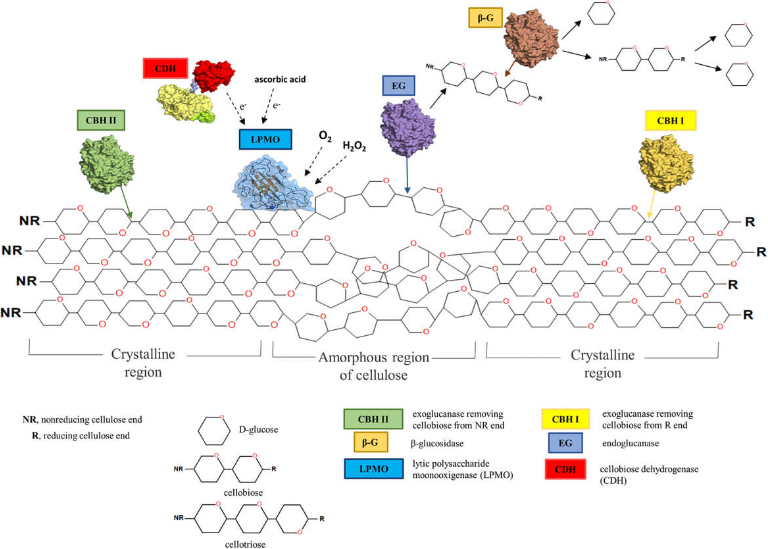
图3、纤维素酶(内切酶、外切酶、β-葡萄糖苷酶)与裂解性多糖单加氧酶(LPMO,如AA9或AA10家族)通过协同作用降解纤维素链的示意图(图片来源:DOI: 10.1002/elsc.201800039)
微生物降解纤维素的分子机制
目前研究较为深入的纤维素降解机制主要有两种:水解机制和氧化机制。此外,褐腐菌还采用一种研究相对较少的独特氧化途径。尽管前两种机制均涉及纤维素酶对β-1,4-糖苷键的断裂,但酶在环境中的呈现方式及作用原理存在显著差异。大多数好氧微生物采用游离纤维素酶机制,而褐腐菌则倾向于利用氧化机制。
A. 水解机制 (Hydrolytic Mechanism)
在糖苷水解酶(Glycosyl hydrolases)家族中,糖苷键的酶促水解通常通过广义酸碱催化实现。该过程需要两个关键残基:质子供体(HA)和亲核试剂/碱(B⁻),通常由天冬氨酸或谷氨酸残基提供。
从机理上看,尽管不同纤维素酶的结构各异,但其活性中心的羧基对均参与广义酸碱催化:
• 一个残基作为广义酸,使糖苷键上的氧原子质子化;
• 另一个残基作为亲核试剂参与反应。
根据两个羧基之间的距离不同,水解机制可分为构型翻转型(Inverting,间距约10 Å)和构型保持型(Retaining,间距约5 Å)。
1. 构型翻转机制 (Inverting Mechanism)
在此机制中,两个酶残基(通常为羧酸盐)分别充当酸和碱。水分子对葡萄糖环的C1碳原子发起SN2型亲核攻击,导致异头碳(C1)的构型发生翻转。
2. 构型保持机制 (Retaining Mechanism)
该机制包含两个连续的步骤,每一步都涉及构型翻转,最终结果表现为构型保持:
• 第一步:亲核试剂攻击异头中心,导致去质子化,形成共价糖基 - 酶中间体。
• 第二步:去质子化的羧酸盐充当碱,协助水分子进行亲核攻击,释放水解产物并再生酶。
3. GH4糖苷酶的特殊机制
近期研究发现,依赖NAD和二价金属离子的GH4家族糖苷酶采用了一种截然不同的机制:
• C3位的氢负离子被夺取生成酮基;
• 随后C2位去质子化,伴随酸催化的糖苷氧消除,形成1,2-不饱和中间体;
• 该α-β不饱和物种经碱催化水合生成3-酮衍生物,最后由NADH还原完成反应循环。
典型代表:水解机制广泛存在于大多数好氧和厌氧微生物中,包括芽孢杆菌属(Bacillus)、不动杆菌属(Acinetobacter)、生孢噬纤维菌属(Cellulomonas)、梭菌属(Clostridium),以及真菌中的黑曲霉(Aspergillus niger)、尖孢镰刀菌(Fusarium oxysporum)、粗糙脉孢菌(Neurospora crassa)和里氏木霉(Trichoderma reesei)。
B. 氧化机制 (Oxidative Mechanism)
尽管大多数好氧细菌依赖多种纤维素酶的协同作用降解纤维素,但某些无细胞真菌培养滤液在有氧条件下的降解速率显著高于厌氧条件。在此过程中,纤维二糖脱氢酶(CDH)发挥了关键作用。
CDH的核心功能:
1. 氧化还原反应:CDH通过“乒乓”机制催化纤维二糖(纤维素酶的主要产物)氧化为纤维二糖内酯,同时还原多种电子受体(如醌类、螯合Fe(III)、O₂生成过氧化氢、苯氧自由基等)。
2. 防止链重缩合:CDH氧化内切纤维素酶产生的游离末端,阻止纤维素链的重新缩合。
3. 解除产物抑制:高浓度的纤维二糖会抑制多种纤维素酶的活性,CDH通过移除纤维二糖有效缓解了这种产物抑制效应。
4. 芬顿反应(Fenton Reaction):CDH能将Fe³⁺还原为Fe²⁺,并将O₂还原为H₂O₂。二者共同引发芬顿反应,产生高活性的羟基自由基(·OH),从而非特异性地解聚或修饰纤维素结构。
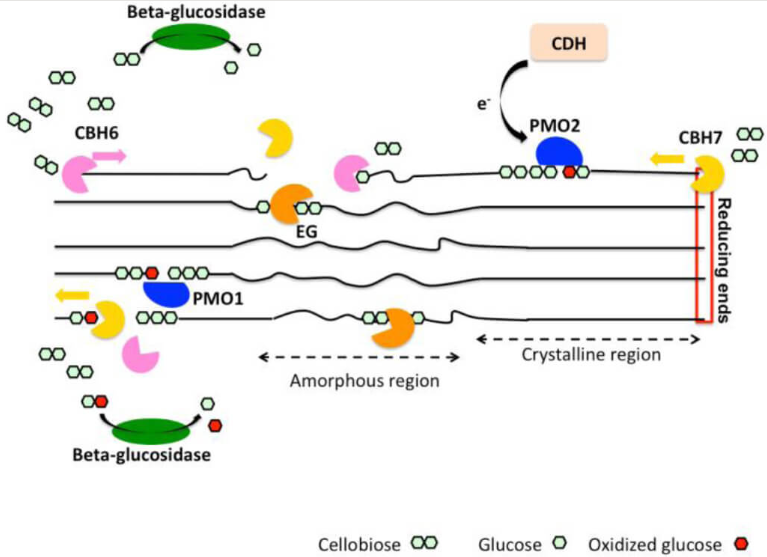
图4、纤维素降解的氧化机制示意图(图片来源:DOI: 10.5936/csbj.201209015)
典型代表:氧化机制主要见于部分真菌物种,如黄孢原毛平革菌(Phanerochaete chrysosporium)、嗜热丝孢菌(Sporotrichum thermophile)、Poria placenta及Lenzites trabea等。
总结
综上所述,微生物对纤维素的降解是一个高度有序、多酶协同且机制复杂的生物化学过程。从内切葡聚糖酶、外切葡聚糖酶到β-葡萄糖苷酶的“三步走”水解策略,
再到褐腐菌等特定类群利用氧化酶系(如CDH)引发的自由基攻击,自然界展现了多样化的降解智慧。
好氧与厌氧环境下的不同代谢路径,以及温度、pH、木质素含量等环境因子的精细调控,共同构成了一个动态平衡的生态系统。
深入理解这些机制不仅揭示了微生物在地球碳循环中的核心地位,更为现代生物技术提供了宝贵的启示。
无论是优化工业酶制剂的配方、设计高效的纤维素酶体复合体,还是开发基于木质纤维素的第二代生物燃料,对降解机理的精准掌握都是技术突破的前提。
未来,随着结构生物学与宏基因组学的进一步融合,我们有望挖掘出更多 novel 酶资源,甚至通过人工设计重构降解路径,从而更高效地将丰富的植物生物质转化为可持续的能源与高附加值化学品,助力全球碳中和目标的实现。
参考文献
https://microbenotes.com/microbial-degradation-of-cellulose/
相关产品
HZB217966:黄孢原毛平革菌 | Phanerochaete chrysosporium
HZB746114:解纤维素醋弧菌ATCC 33288 | Acetivibrio cellulolyticus (CD2)
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-03-20
编制人:小灰
审稿人:小藻