半纤维素微生物降解的酶系、机制与调控全景
来源:武汉市灰藻生物科技有限公司 浏览量:254 发布时间:2026-03-30 16:23:47
引言
作为植物细胞壁的三大核心组分之一的半纤维素,其结构复杂且高度异质,长期以来被视为生物质精炼与全球碳循环中的“顽固堡垒”。
不同于结晶度高的纤维素,半纤维素由多种戊糖、己糖及糖醛酸通过复杂的糖苷键连接而成,并带有丰富的侧链修饰,这种独特的无定形结构既赋予了植物细胞壁柔韧性,也为其生物降解带来了巨大挑战。
在自然界的物质循环中,微生物凭借其分泌的多样化半纤维素酶系,成为了破解这一复杂多糖网络的关键力量。
从森林土壤到反刍动物瘤胃,再到海洋生态系统,微生物通过外切与内切酶的精密协同,高效地将半纤维素转化为可被利用的单糖与寡糖。
本文旨在系统梳理半纤维素的化学结构特征,深入剖析参与降解过程的各类微生物资源(真菌与细菌),详细阐述四大类关键酶系(木聚糖酶、甘露聚糖酶、半乳聚糖酶及阿拉伯聚糖酶)的作用机制,并探讨温度、pH及底物特性等环境因子对降解效率的调控规律。通过全方位解析这一生物转化过程,我们期望为生物质能源开发、农业废弃物资源化利用以及新型酶制剂的研发提供坚实的理论支撑。
何为半纤维素?
半纤维素是一类复杂的杂聚多糖,广泛存在于植物纤维中,常与纤维素、果胶等多糖共存。
不同于结构均一的纤维素(由葡萄糖单体组成的同聚物),半纤维素是由多种碳水化合物构成的异质性群体,其具体结构迄今尚未完全阐明。各类半纤维素在结构与理化性质上差异显著:它们与纤维素无结构关联,单体组成也不尽相同,但部分可溶于水或碱液。除阿拉伯半乳聚糖外,大多数半纤维素具有较短的侧链和较低的分子量。
半纤维素的单体组成丰富,既包含戊糖(如木糖、阿拉伯糖),也包含己糖(如葡萄糖、甘露糖、半乳糖)以及糖醛酸。这些多糖在植物体内兼具吸水、储能及支撑功能。
尽管“半纤维素(hemicellulose)”这一术语略显陈旧,且学界曾提议以“交联聚糖(cross-linking glycans)”取而代之,但由于“交联”并非所有半纤维素的普遍特征,该替代名称尚存争议。因此,“半纤维素”一词至今仍被广泛沿用。
根据主链单糖残基的不同,半纤维素主要分为以下几类:
木葡聚糖(xyloglucan)、木聚糖(xylans)、甘露聚糖(mannans)、葡甘露聚糖(glucomannans),以及β-(1→3, 1→4)-葡聚糖。
这些聚糖在C1和C4位均具有相同的平伏键构型,因而其主链结构表现出显著的相似性。其中,木聚糖是该类多糖的代表,由β-(1→4)连接的木糖残基构成主链。作为自然界中含量仅次于纤维素的多糖,木聚糖的研究价值不言而喻。
半纤维素的结构特征
与纤维素聚合物中包含7000至15000个葡萄糖分子不同,半纤维素的聚合度较低,通常由50至3000个糖单元组成。
结构上,半纤维素呈无定形态,而非纤维素的结晶态,这使得其更易受热液提取和水解作用的影响。依据主链主要糖残基的类型,半纤维素可进一步细分为木聚糖、甘露聚糖和葡聚糖等类别。
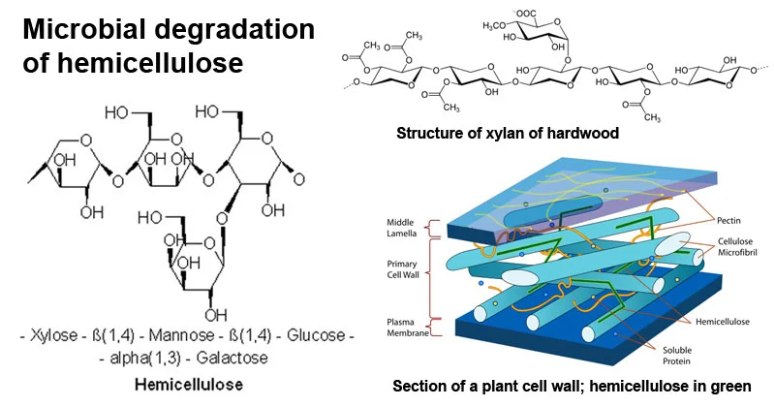
图1、半纤维素的结构特征(图片来源:Wikipedia)
1. 木聚糖 (Xylans)
木聚糖是一类由β-(1→4)连接的木糖残基构成主链,并带有α-阿拉伯呋喃糖和α-葡萄糖醛酸侧链的多糖,这些侧链有助于其与纤维素的交联。
木聚糖可分为三类:葡萄糖醛酸木聚糖、阿拉伯木聚糖以及葡萄糖醛酸阿拉伯木聚糖。
木聚糖缺乏重复的结构单元,其结构的多样性至今仍未被完全解析。
木聚糖的一个共同特征是,其O-2位连接的木糖残基常作为阿魏酰阿拉伯呋喃糖侧链的取代基存在。
大多数木聚糖存在不同程度的乙酰化修饰,尤其在双子叶植物的次生壁中更为显著。乙酰基团主要连接在木糖残基的O-3位,较少部分连接在O-2位。
2. 甘露聚糖 (Mannans)
甘露聚糖是一类主链完全由甘露糖单元通过β-(1→4)键连接而成的多糖。
由于其作为种子储存物质的重要角色,甘露聚糖已被广泛研究,但它们也以不同含量存在于所有细胞壁中。
值得注意的是,酵母中的甘露聚糖结构较为特殊,其主链由α-(1→6)键连接,并带有α-(1→2)和α-(1→3)键连接的支链。
3. 葡聚糖 (Glucans)
葡聚糖是由葡萄糖单元通过糖苷键连接而成的多糖。在半纤维素中,葡聚糖主要表现为木葡聚糖或β-(1→3,1→4)-葡聚糖。
• 木葡聚糖:由β-(1→4)连接的葡聚糖主链构成,并取代有木糖分子。
• β-(1→3,1→4)-葡聚糖:这类混合连接葡聚糖以β-(1→4)连接为主,其间穿插单个β-(1→3)连接键。其结构主要由通过β-(1→3)键连接的纤维三糖基和纤维四糖基单元主导,但也存在较长的β-(1→4)连接片段。
什么是半纤维素酶?
半纤维素酶是一类专一性降解半纤维素的酶系。除了作用于糖苷键外,它们还能有效水解半纤维素主链上的短链或单糖侧链。
鉴于半纤维素结构的复杂性与多样性,其完全降解往往需要多种酶的协同作用。
半纤维素酶的催化模块主要分为两类:
• 糖苷水解酶 (GHs):负责水解糖苷键;
• 碳水化合物酯酶 (CEs):负责水解乙酸或阿魏酸侧基的酯键。
作为自然废物循环系统的关键组成部分,半纤维素酶在全球碳循环中扮演着至关重要的角色。
根据作用底物和化学键类型的不同,半纤维素酶可分为四大类:L-阿拉伯聚糖酶、D-半乳聚糖酶、D-甘露聚糖酶和D-木聚糖酶。
这些酶不仅在结构上存在差异,其作用模式也各不相同。此外,细菌、真菌、原生动物、藻类乃至植物等不同生物类群均可产生不同类型的半纤维素酶。
参与半纤维素降解的微生物
1. 半纤维素降解真菌
真菌是自然界中分解有机物质最活跃的类群之一,因此也是最重要的半纤维素降解微生物。
好氧真菌(如木霉属 Trichoderma 和曲霉属 Aspergillus)能分泌高浓度的多种半纤维素酶,这些酶之间具有显著的协同效应。
半纤维素酶的来源取决于微生物所产生的具体酶系类型。
此外,多种腐生真菌也被证实能产生 L-阿拉伯聚糖酶和 D-甘露聚糖酶,包括链格孢 (Alternaria solani)、葡萄座腔菌 (Botryosphaeria ribis)、葱灰葡萄孢 (Botrytis allii)、离心革裢菌 (Corticium centrifugum)、果生单端孢 (Monilia fructigena)、脉孢菌 (Neurospora)、指状青霉 (Penicillium digitatum)、黑根霉 (Rhizopus nigricans) 以及核盘菌 (Sclerotinia fructigena) 等。
同样, Gibberella saubinetti、稻平脐蠕孢 (Helminthosporium oryzae)、致病疫霉 (Phytophthora infestans)、Gibbosa 栓菌 (Trametes gibbosa) 等真菌则能产生 D-半乳聚糖酶。
海洋环境中的真菌物种,如 sojae 曲霉 (Aspergillus sojae)、球毛壳菌 (Chaetomium globosum)、双孢蘑菇 (Agaricus bisporus)、葡萄座 Diplodia (Diplodia viticola)、Oxiporus 属等,也被视为半纤维素酶的重要来源。
大多数半纤维素酶为胞外酶,但也存在部分胞内半纤维素酶。2. 半纤维素降解细菌
细菌也能产生不同类型的半纤维素酶,既可以是单一酶,也可以是多酶复合体系。
目前已报道的产纤维素酶细菌主要包括:溶纤维梭菌 (Clostridium felsineum)、枯草芽孢杆菌 (Bacillus subtilis)、解甘露糖乙酰杆菌 (Acetenobacter mannanolyticus)、AROIDEAE 芽孢杆菌 (Bacillus aroideae) 以及粘球菌属 (Sporocytophaga myxococcoides) 等。
此外,存在于各类反刍动物瘤胃中的细菌也被证实能产生半纤维素酶,典型代表包括链球菌属 (Streptococcus sp.)、溶纤维丁酸弧菌 (Butyrivibrio fibrisolvens)、白色瘤胃球菌 (Ruminococcus albus) 以及瘤胃拟杆菌 (Bacteroides ruminicola) 等。
参与半纤维素降解的关键酶系
半纤维素酶通常分为四大类:L-阿拉伯聚糖酶、D-半乳聚糖酶、D-甘露聚糖酶和 D-木聚糖酶。
1. L-阿拉伯聚糖酶 (L-arabinanases)
L-阿拉伯聚糖酶是一类水解酶,能够降解 L-阿拉伯聚糖,其作用位点包括 L-阿拉伯聚糖上的 α-(1→3) 连接的 L-阿拉伯呋喃糖侧链 appendages,以及“线性”主链上的 α-(1→5) 连接的 L-阿拉伯呋喃糖残基。
据报道,溶纤维梭菌西科基亚变种 (Clostridium felsineum var. sikokianum)、腐生及植物病原真菌、蜗牛以及植物均能产生此类酶。
瘤胃细菌、原生动物以及盲肠细菌也能产生可从阿拉伯木聚糖中释放 L-阿拉伯糖的酶,这类酶可能属于 α-L-阿拉伯呋喃糖苷酶。
大多数真菌源性的 L-阿拉伯聚糖酶通常被分泌到培养基中(胞外),但也发现了胞内 L-阿拉伯聚糖酶的存在。
已知多种植物病原真菌在含有 L-阿拉伯聚糖的培养基中生长时,会通过诱导作用产生 L-阿拉伯聚糖酶;而当以 D-葡萄糖为唯一碳源时,这些生物则会组成型地产生该酶。
2. D-半乳聚糖酶 (D-galactanases)
D-半乳聚糖酶是一类能够降解 D-半乳聚糖和 L-阿拉伯-D-半乳聚糖的水解酶。
目前已发现两种 distinct 类型的 D-半乳聚糖酶,分别特异性作用于 (1→3) 和 (1→4) 的 β-D-吡喃半乳糖苷键。
这两种酶均能随机降解 D-半乳聚糖,生成 D-半乳糖和 D-半乳寡糖,因此属于内切-D-半乳聚糖酶 (endo-D-galactanases)。
据报道,枯草芽孢杆菌、瘤胃厌氧细菌、真菌以及植物均能产生 D-半乳聚糖酶。
D-半乳聚糖酶的合成具有诱导性,微生物源性的该类酶通常响应培养基中的碳源而被诱导并在胞外分泌。
3. D-甘露聚糖酶 (D-Mannanases)
D-甘露聚糖酶 [即 (1→4)-β-D-甘露聚糖甘露糖水解酶,或称内切-D-甘露聚糖酶] 是一类能够水解 D-甘露聚糖和 D-半乳-D-甘露聚糖中 (1→4)-β-D-吡喃甘露糖苷键的水解酶。
此类酶能够降解 D-葡-D-甘露聚糖,生成 D-葡萄糖、D-甘露糖以及一系列甘露寡糖和葡甘露寡糖。
据报道,多种细菌(包括来自人类肠道和瘤胃的细菌)均能产生 D-甘露聚糖酶。
其他来源还包括瘤胃原生动物、各类真菌(涵盖腐生菌、植物病原菌及菌根真菌)、海洋藻类、萌发的陆生植物种子以及多种无脊椎动物。
4. β-D-木聚糖酶 (β-D-xylanases)
β-D-木聚糖酶是一种水解酶,能将 β-D-木聚糖降解为 D-木糖或 D-木糖与 D-木寡糖的混合物。
木聚糖酶主要分为两类,分别降解 (1→3) 键和 (1→4) 键。
D-木聚糖酶广泛分布于海洋和陆地环境的细菌、真菌(腐生菌、植物病原菌及菌根真菌)、瘤胃细菌和原生动物、反刍动物盲肠细菌、昆虫、蜗牛、甲壳类动物、海洋藻类以及陆生植物的萌发种子中。
影响半纤维素降解的因素
半纤维素的降解效率受多种因素影响,主要包括以下几点:
1. 温度与 pH 值
温度和 pH 值均显著影响半纤维素的水解速率。实验表明,在 pH 6.0 和 40°C 条件下,水解速率达到峰值。
当反应温度超过 40°C 时,总木糖产量反而下降,这可能是因为高温影响了半纤维素酶的热稳定性。
2. 有机质含量
有机质的存在通常会加速半纤维素的降解,因为大量有机质可作为微生物的底物。
然而,若体系中仅含半纤维素而无其他组分,水解速率则会降低。
添加少量易分解的有机质可促进微生物生长,从而显著提高降解速率。
3. 酶投加量
增加酶浓度可直接提升半纤维素的降解速率。
酶浓度的提高增加了活性位点的数量,进而提升了产物浓度。
4. 底物转化特性
在酶水解反应进行数小时后,木糖和阿拉伯糖的生成速率会迅速下降。
值得注意的是,水解过程中释放的阿拉伯糖比例始终显著低于同期产生的木糖比例。
水解速率的急剧下降,主要归因于随着水解的进行,细胞壁基质结构对酶的可达性(accessibility)逐渐受限,而非酶本身的性质发生了改变。
降解过程简述
半纤维素的降解过程与纤维素等同聚物的降解大体相似,但为了获得单体单元,半纤维素降解通常遵循以下两条路径之一:
1. 侧链去除路径:降解始于外切糖苷酶(exoglycosidases)对半纤维素的攻击,旨在移除侧链取代基。这一步骤相当于“打开”或暴露出聚糖主链,显著降低了侧链残基造成的空间位阻(steric hindrance),从而使主链更易受到半纤维素酶的攻击。
2. 主链切断路径:降解始于内切半纤维素酶(endohemicellulases)直接攻击聚糖链上无分支或分支较少的区域。随后,内切酶的作用产生一系列组成混合的寡糖片段。最终,外切糖苷酶和内切半纤维素酶协同作用,将这些片段进一步降解。
半纤维素微生物降解的机制
微生物降解的具体模式或机制,通常只有在所使用的酶制剂为均一组分(即单一蛋白成分)时才能被清晰阐释。不同类型的半纤维素酶,其作用机制各异。
1. 木聚糖酶 (Xylanases)
目前表征最为清楚的是内切型 D-木聚糖酶。
• 作用底物:这类酶水解 D-聚糖(如 L-阿拉伯-D-木聚糖、L-阿拉伯-D-葡萄糖醛酸-D-木聚糖及 D-葡萄糖醛酸-D-木聚糖)中的 1,4-β-D-吡喃木糖苷键。
• 特殊功能:部分酶甚至能水解阿拉伯木聚糖中的 (1→3)-α-L-阿拉伯呋喃糖分支点。
• 产物:另一类内切木聚糖酶可将阿拉伯木聚糖及其他 D-木聚糖降解为 D-木糖、D-木寡糖,在某些情况下还能产生同时含有 L-阿拉伯糖和 D-木糖的寡糖。
实例:细菌木聚糖酶
芽孢杆菌属 (Bacillus) 和链霉菌属 (Streptomyces) 等细菌可产生木聚糖酶。例如,来自嗜碱芽孢杆菌的木聚糖酶制剂,能将阿拉伯木聚糖主要降解为木二糖和木三糖,并伴随少量高分子量的木寡糖。
2. 甘露聚糖酶 (Mannanases)
已表征的甘露聚糖酶包括外切型和内切型,它们均能水解支链甘露聚糖、共聚甘露聚糖及线性 D-甘露聚糖中的 1,4-β-D-吡喃甘露糖苷键。
• 内切-β-甘露聚糖酶:将 β-D-甘露聚糖降解为 D-甘露糖及一系列甘露寡糖。若结合 β-D-甘露糖苷酶进行酸水解,D-甘露糖将是唯一的水解产物。
• 优先攻击位点:内切甘露聚糖酶优先攻击分子非还原端起的第 3 和第 4 个连接键处的 D-甘露糖链。
实例:真菌甘露聚糖酶
真菌源性的 D-甘露聚糖酶已知以随机方式降解 D-甘露聚糖,且属于内切型。由半乳葡甘露聚糖酶解产生的混合寡糖中,除 D-葡萄糖和 D-甘露糖外,通常还含有 D-半乳糖。
3. 半乳聚糖酶 (Galactanases)
半乳聚糖酶是一类水解酶,能够降解 D-半乳聚糖和 L-阿拉伯-D-半乳聚糖。
目前已识别出两种 distinct 类型的内切半乳聚糖酶和一种外切型。
• 作用机制:内切半乳聚糖酶随机降解 D-半聚糖中的 1,4-β-D-半乳糖苷键,生成 D-半乳糖和半乳寡糖,部分产物中可能含有 L-阿拉伯糖残基。
实例:真菌半乳聚糖酶
根霉属 (Rhizopus sp.) 产生的 D-半乳聚糖酶对半乳二糖无作用,因其特异性仅针对 (1→3)-β-D-吡喃半乳糖苷键。此外,该类酶能从阿拉伯半乳糖苷中移除 L-阿拉伯呋喃糖,但无法从寡糖中释放游离的 L-阿拉伯糖。
4. 阿拉伯聚糖酶 (Arabinanases)
• 内切型作用:作为内切酶,L-阿拉伯聚糖酶水解的主要产物是 L-阿拉伯糖和 L-阿拉伯寡糖,同时也可能生成少量 L-阿拉伯二糖。
• 外切型作用:外切型 L-阿拉伯聚糖降解酶能将 L-阿拉伯聚糖完全降解为 L-阿拉伯糖。
• 双重水解能力:这些酶在单一活性位点上即可水解 L-阿拉伯聚糖中的 (1→3) 和 (1→5) 两种 α-L-阿拉伯呋喃糖残基连接键。底物通过多链机制从非还原端受到攻击。
• 动力学特征:在攻击 L-阿拉伯聚糖时,酶首先快速水解底物约 30%;随后反应速率显著减慢。
o 机理解析:初期的快速水解对应于酶优先攻击 (1→3)-α-L-阿拉伯呋喃糖残基;留下的主要是线性的 (1→5)-α-L-阿拉伯聚糖主链,这部分随后被缓慢但最终完全地水解为 L-阿拉伯糖。
结语
综上所述,半纤维素的微生物降解是一个高度复杂且精密的生物化学过程,依赖于微生物群落与多功能酶系的动态协同。从去除侧链取代基以消除空间位阻,到内切酶随机切断主链生成寡糖片段,再到外切酶进一步水解为单体,这一系列步骤展现了自然界在分子水平上的精妙设计。
研究表明,无论是好氧真菌还是厌氧细菌,亦或是瘤胃共生微生物,它们所分泌的半纤维素酶系(GHs与CEs)均表现出显著的底物特异性和环境适应性。然而,降解效率并非一成不变,而是受到温度、pH、有机质含量及酶投加量等多重因素的严格调控。特别是底物结构的物理可达性(accessibility)往往成为限制水解速率的关键瓶颈。
展望未来,随着合成生物学与蛋白质工程技术的进步,深入理解半纤维素降解的微观机制将有助于我们构建更高效的人工酶系组合,优化生物炼制工艺。这不仅对于推动第二代生物燃料的产业化进程具有重要意义,更是实现农业废弃物高值化利用、促进全球碳循环平衡的关键所在。解开半纤维素的“密码”,将为绿色生物制造开启无限可能。
参考文献
https://microbenotes.com/microbial-degradation-of-hemicellulose/
相关产品
HZB122445:茄链格孢(番茄早疫病菌) | Alternaria solani (IVF094)
HZB912093:溶纤维丁酸弧菌CCUG 35459T | Butyrivibrio fibrisolvens (DSM 3071)
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-03-25
编制人:小灰
审稿人:小藻