细胞的“时间暂停”魔法:微生物超低温冻存与冷冻干燥的底层科学机制
来源:武汉市灰藻生物科技有限公司 浏览量:424 发布时间:2026-03-25 20:51:08
一、引言
在生命科学研究与工业发酵的宏大蓝图中,微生物细胞工厂是真正的“核心芯片”。
然而,自然界中的微生物为了适应环境,其基因组始终处于动态突变之中。无论是进行极其严密的生长动力学测定、代谢物靶向收集,还是规模化的生物反应器放大,
如果作为起点的菌株发生了质粒丢失,或关键代谢表型的退化,所有下游数据都将失去科学价值。
为了对抗这种由时间带来的熵增,科学家们开发了超低温冻存(Cryopreservation)与冷冻干燥(Lyophilization,简称冻干)技术。
这并非简单的物理降温或水分抽干,而是一场在微米级尺度上进行的精妙“生化干预”。
本文将跨越表象,从细胞生物物理学、热力学相变机制,以及复苏后的生长动力学验证等多个维度,深度拆解这两种“时间暂停魔法”背后的科学原理,揭示高质量菌种保藏的技术壁垒。
二、超低温冻存的物理学博弈:两因素损伤假说
超低温冻存通常在 -80°C 的超低温冰箱或 -196°C 的液氮中进行。在液氮温度下,细胞内的一切酶促反应、DNA复制和生化代谢都会因为缺乏足够的热力学能量而陷入绝对的停滞。
然而,降温的过程本身却充满了致命的危险。
2.1 溶质效应与胞内冰晶的双重绞杀
著名的冷冻生物学家 Peter Mazur 提出了经典的“两因素损伤假说(Two-Factor Hypothesis)”,完美解释了细胞在冷冻过程中面临的生死考验:
降温过慢导致的“溶质效应(Solution Effects)”:当降温较慢时,胞外的自由水率先结冰,导致未结冰的胞外溶液浓度急剧飙升。
此时,细胞内的大量水分因强烈的渗透压差被迫外流。细胞发生严重脱水皱缩,高浓度的胞内电解质会直接破坏蛋白质结构并溶解细胞膜的脂质双分子层。
降温过快导致的“胞内冰晶(Intracellular Ice Formation)”:如果降温极其猛烈,胞内的水分子根本来不及渗出细胞,就会在原位迅速冻结。
形成的长针状冰晶如同微观的匕首,会无情地刺破细胞膜、内质网乃至染色体,造成细胞不可逆的机械死亡。
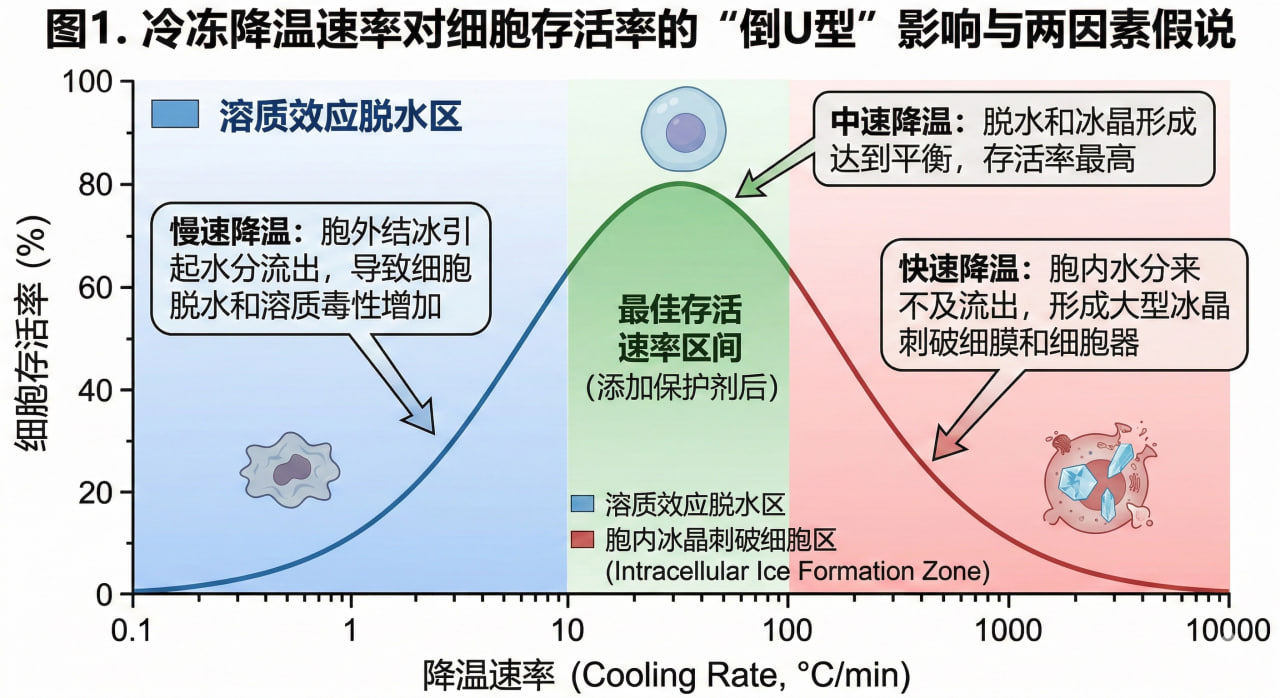
- 图1. 冷冻降温速率对细胞存活率的“倒U型”影响与两因素假说
2.2 渗透性保护剂与“玻璃化(Vitrification)”状态
为了在慢速脱水与快速结冰之间找到一条生路,必须引入渗透性冷冻保护剂(Penetrating Cryoprotectants),如甘油(Glycerol)和二甲基亚砜(DMSO)。
这些小分子物质不仅能迅速穿透细胞膜进入胞内,降低溶液的冰点,更重要的是,它们极强的水合能力能够与水分子形成密集的氢键网络。
在超低温下,这些保护剂能迫使胞内液体绕过结晶相,直接进入一种高粘度的非晶态固体结构,即玻璃化(Vitrification)状态。
在玻璃化状态下,水分子被“冻结”在混乱排列的瞬间,彻底避免了尖锐冰晶的产生,从而完美地保留了细胞的完整形态。
三、冷冻干燥的真空升华奇迹:极限脱水的艺术
尽管液氮冻存是维持遗传稳定性的黄金标准,但其对冷链物流和持续供能的严苛要求,限制了微生物在全球范围内的共享与运输。
冷冻干燥技术(Lyophilization)通过物理学上的“升华”现象,实现了菌种在常温条件下的长效保藏。
3.1 热力学三相图与升华控制
冻干的核心在于精准操控水的三相态。其工艺过程极其严密:
预冻结冰,寻找共晶点(Eutectic Point):首先必须将菌液降温至完全冻结,特别是要低于整个混合溶液的共晶点温度,
确保没有一丝液态游离水存在,否则在后续抽真空时会发生剧烈的沸腾“喷瓶”现象。
一次干燥(升华解析):在深度真空(低于三相点气压)环境下,温和地输入热量。
此时,固态的冰直接升华成水蒸气被冷阱捕获,去除了细胞内约 90% 的宏观水分。
二次干燥(解吸附):继续升高搁板温度,利用分子热运动的加剧,强行剥离与蛋白质、
核酸等生物大分子通过氢键紧密相连的“结合水”,最终使残余水分降至 1%~3% 的安全区间。
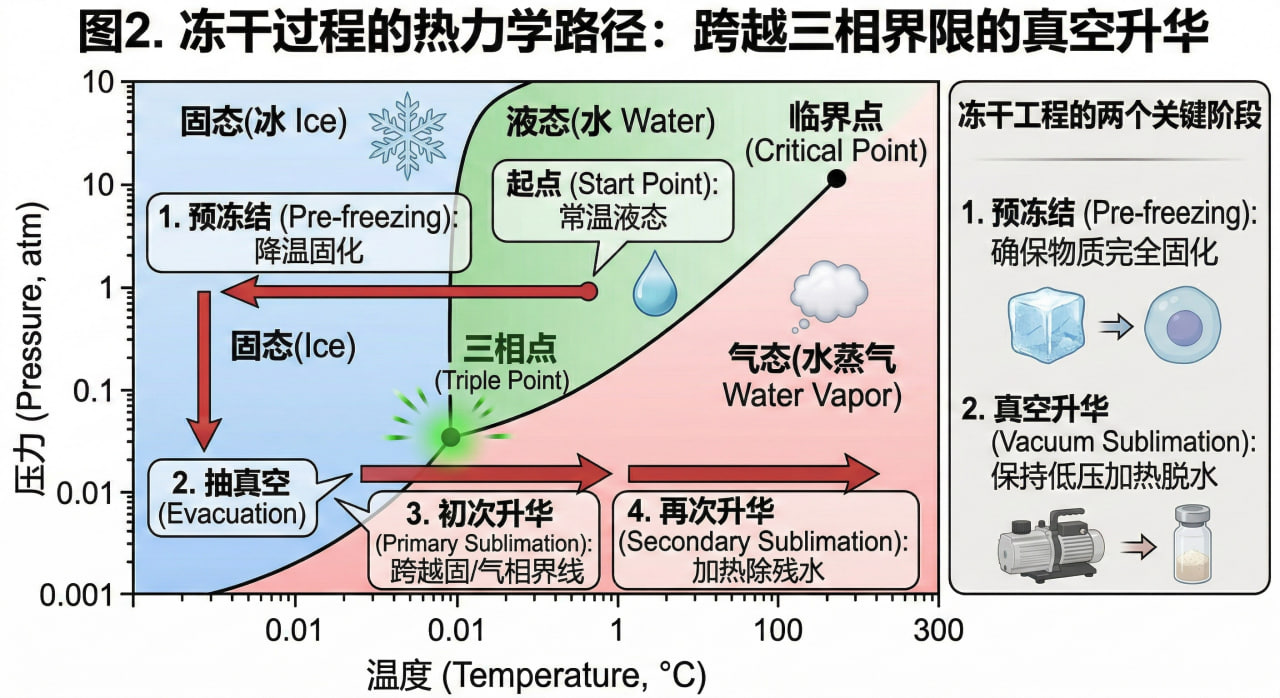
图2. 冻干过程的热力学路径:跨越三相界限的真空升华
3.2 非渗透性保护剂:海藻糖的“水替代假说”
细胞膜的磷脂双分子层高度依赖水合层来维持流动性。一旦失去“结合水”,脂质体会发生致命的相变破裂。
此时,非渗透性保护剂(如海藻糖 Trehalose、蔗糖或脱脂乳粉)成为了生命最后的护盾。
根据公认的“水替代假说(Water Replacement Hypothesis)”,海藻糖分子上的大量羟基能够完美嵌合到脱水磷脂的亲水端,替代原本游离水的位置,维持了磷脂分子的空间间距。
结合“玻璃态动力学假说”,这些保护剂分子在细胞外部形成了一层坚不可摧的非晶态糖玻璃骨架,将脆弱的蛋白质包裹其中,防止了脱水导致的蛋白质三级结构崩塌与聚集变性。
四、特殊挑战:厌氧菌的保藏与氧化应激
常规的好氧细菌(如枯草芽孢杆菌、大肠杆菌)保藏相对简单,但对于从事特殊代谢物研究的实验室而言,
严格厌氧菌(Strict Anaerobes)(如双歧杆菌、梭菌属等)的保藏是一项极具挑战性的技术壁垒。
厌氧菌极其缺乏过氧化氢酶或超氧化物歧化酶(SOD)。在制备冻干保护液、分装安瓿瓶乃至复苏的短暂瞬间,只要接触到微量氧气,细胞内就会迅速生成具有极强破坏性的活性氧(ROS)。
ROS 会像猛兽一样撕裂厌氧菌的核酸并氧化细胞膜脂质。
因此,高标准的厌氧菌保藏必须在严格的无氧手套箱中进行,保护剂中往往需要添加抗坏血酸、半胱氨酸等强效还原剂作为“氧气清道夫”,
并使用刃天青(Resazurin)作为氧化还原电位的实时监控指示剂,确保细胞在休眠全程免受氧化应激损伤。
五、复苏的科学:生长动力学与延滞期验证
无论冻存技术多么精妙,保藏的终极目的都是为了能够完美复苏并重现其生理特征。唤醒休眠的细胞,是一场精密的生化重启过程。
5.1 渗透压缓冲与损伤修复
处于高度脱水状态的冻干菌粉,在接触液体的瞬间面临着极大的渗透压冲击。
标准操作中,必须使用富含缓冲盐和营养素的液体恢复培养基进行缓慢滴加水化,给细胞膜脂双层一个“重塑”的时间窗口。
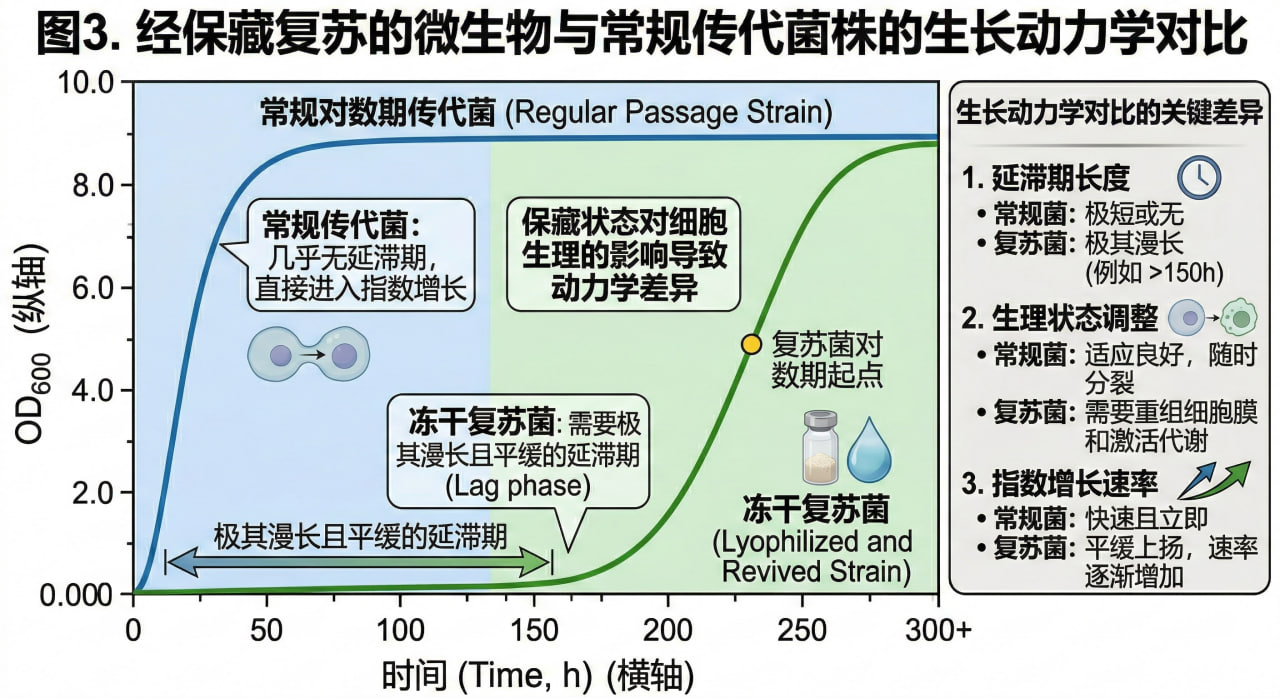
图3. 经保藏复苏的微生物与常规传代菌株的生长动力学对比
5.2 绘制标准化生长曲线与表型确认
刚复苏的细胞并不会立刻进行分裂增殖。在分光光度计(OD600)监测的生长曲线中,研究者会观察到复苏菌株存在一段异常漫长的延滞期(Lag phase)。
在这一阶段,细胞正疯狂调动残存的能量,进行因冷冻损伤导致的 DNA 链断裂修复、核糖体的重新组装以及逆境蛋白的降解。
对于严谨的实验室而言,仅仅能“长出菌落”是不够的。必须将复苏后的细胞接种入新鲜培养基,进行密集的取样,绘制出至少三个生物学重复的标准化生长曲线,并同步收集其代谢产物。
只有当复苏菌株的世代时间(Generation time)、最大比生长速率(μmax)以及目标代谢物产量与建库时的初始数据高度重合,这一次的“时间暂停”魔法才算真正宣告成功。
六、结语与技术展望
从 -196°C 极寒中的玻璃化静止,到极限真空下的升华脱水,微生物超低温保藏与冷冻干燥技术,是物理化学与细胞生物学完美交融的结晶。
随着合成生物学的爆发,我们面对的不再是简单的野生菌株,而是基因组经过大规模重编、代谢网络高度脆弱的“定制化超级细胞工厂”。
优化更加智能的复合型冷冻保护剂,探索更加精准的冻干升华曲线控制,是保障生物制造产业根基的重中之重。以科学为盾,对抗时间的侵蚀,这是每一位生物科技工作者的神圣使命。
参考文献
1. Mazur, P. (1984). Freezing of living cells: mechanisms and implications. American Journal of Physiology-Cell Physiology, 247(3), C125-C142.
2. Crowe, J. H., et al. (1992). The role of vitrification in anhydrobiosis. Annual Review of Physiology, 60, 73-103.
3. Morgan, C. A., et al. (2006). Preservation of micro-organisms by drying; a review. Journal of Microbiological Methods, 66(2), 183-193.
相关技术与支撑体系
HZB208299:EBY100 酿酒酵母 | Saccharomyces cerevisiae EBY100 (采用优化甘油/海藻糖复合保护体系)
{kind=link}
{kind=link}
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-03-26
编制人:思琪
审稿人:小藻