关于微生物在生物修复中的作用的系统性综述
来源:武汉市灰藻生物科技有限公司 浏览量:212 发布时间:2026-03-25 20:57:20
摘要
全球环境污染及其修复是亟待解决的重大难题。重金属、染料、碳氢化合物及农药等是导致环境恶化的主因,而残留污染物的清除极具挑战。
生物修复(Bioremediation)利用细菌、真菌和植物降解、去除、转化或固定环境废物,被视为恢复受损生态最成功的方法之一。
该技术通过生物强化(Bioaugmentation)、生物刺激(Biostimulation)、生物通风(Bioventing)等手段提升微生物效能,甚至利用基因工程微生物(GEMs)加速毒素分解。
本文综述了生物修复的最新进展、微生物降解机制及其作为全球污染缓解策略的应用前景。
一、引言
人口激增、城市化及工业化导致大量废物排放,引发严重的环境污染。
污染物涵盖重金属(Cd, Pb, Hg, Cu)、挥发性有机化合物(VOCs)、无机盐、染料、石油烃及多氯联苯(PCBs)等,广泛波及土壤、水体及生态系统。
相比传统物理化学法(成本高、易造成二次污染),生物修复凭借经济、环保及高效的优势成为首选。虽然植物也可用于修复,但微生物因易于控制且生长周期短而更受青睐。
微生物通过酶促或代谢活动(氧化、解毒等)消除污染物。根据实施地点,生物修复分为原位(In situ)和异位(Ex situ)两类。
本文旨在阐述微生物在生物修复中的应用现状,为评估修复过程提供理论依据。
二、生物修复的类型
生物修复过程中采用了多种处理技术或程序(见图1)。
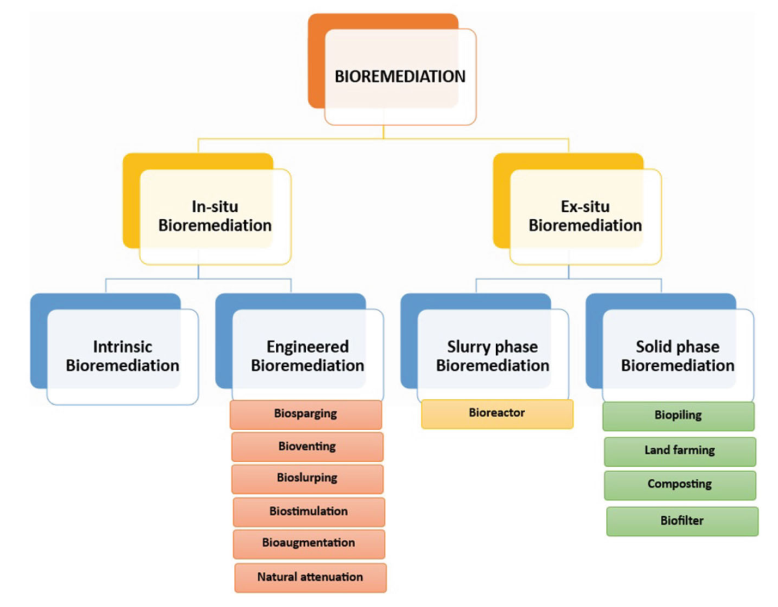
图1.生物修复的类型
2.1 原位生物修复 (In-Situ Bioremediation)
指在污染现场直接处理,无需挖掘转移。其选择取决于场地条件、成本及污染特征。相比之下,异位修复虽需挖掘转移,但更易控制且处理速度更快。
- 内在生物修复 (Intrinsic Bioremediation):又称自然衰减,利用土著微生物在无人为干预下被动降解污染物,成本最低。
- 工程化原位修复(Engineered In-Situ):通过引入特定微生物或优化环境条件加速降解。主要技术包括:
2.2 异位生物修复 (Ex-Situ Bioremediation)
指将污染物挖掘转移至他处处理,适用于污染程度高或需快速修复的场景。
- 泥浆相修复 (Slurry Phase):在生物反应器(Bioreactor)中将污染土壤与水、营养物混合成泥浆,创造理想降解环境。
- 此法速度快,但需后续固液分离处理。反应器类型多样,包括搅拌罐式、气升式及流化床式等。
- 固相修复(Solid Phase):将污染土壤堆置处理,主要包括:
三、生物修复中的微生物
3.1 好氧修复 (Aerobic)
在有氧环境下,多种微生物可修复不同类型的环境污染物。许多复杂的有机分子可被好氧细菌分解,常见的菌属包括鞘氨醇单胞菌属(Sphingomonas)、
红球菌属(Rhodococcus)、芽孢杆菌属(Bacillus)、黄杆菌属(Flavobacterium)、分枝杆菌属(Mycobacterium)、诺卡氏菌属(Nocardia)和假单胞菌属(Pseudomonas)。
研究证实,这些细菌能降解多环芳香化合物、烷烃和农药。许多此类微生物直接从这些污染物中获取能量和碳源。
然而,在有氧生物修复过程中,氧气的供应量往往是限制微生物生长和降解效率的关键因素。
3.2 厌氧修复 (Anaerobic)
研究人员正利用兼性厌氧细菌进行氯化溶剂(如三氯乙烯和氯仿)、多氯联苯及氯代化合物的生物修复。
这些微生物能将污染物降解并转化为毒性较低的形式。厌氧生物修复技术已应用的细菌包括硫酸盐还原菌、气单胞菌属(Aeromonas)和假单胞菌属(Pseudomonas)等。
在不同环境条件下,微生物可对偶氮染料进行降解。利用有机底物氧化产生的电子进行还原反应,可实现偶氮染料的厌氧分解。
由于这些受控的脱色过程,微生物的电化学特性会显著影响脱色效率。出于商业目的,人们利用厌氧脱色产生了随时间变化的脱色代谢物(DMs)。
为了构建更精确的系统,研究者还对特定条件下收集的 DMs 进行了外部强化,以开展更深入的研究(见表1)。
表1.微生物在生物修复中的作用
| 序号 | 污染物 | 物种 | 推论/结论 | 参考文献 |
|---|---|---|---|---|
| 细菌 (Bacteria) | ||||
| 1 | 萘 - 多环芳烃 (PAH) 和重金属 | Bacillus licheniformis(地衣芽孢杆菌)和 Bacillus sonorensis(索诺拉芽孢杆菌)菌株 | PAH 和重金属的生物修复 | Rabani et al. (2022) |
| 2 | 溢油(碳氢化合物) | Azospirillum(固氮螺菌属)、Pseudoalcaligenes(假碱杆菌属)、Acinetobacter(不动杆菌属)、Alcaligenes(产碱杆菌属)、Pseudomonas(假单胞菌属)、Pseudomonas stutzeri(斯氏假单胞菌)和 Paenibacillus(类芽孢杆菌属)物种 | 溢油的修复 | Machado et al. (2019) |
| 3 | 苯、甲苯、乙苯、二甲苯异构体 | Pseudomonas putida(恶臭假单胞菌)、Pseudomonas synxantha(黄绿假单胞菌)和 Pseudomonas azotoformans(生氮假单胞菌) | 二甲苯异构体的降解 | Miri et al. (2022) |
| 4 | 碳氢化合物和多环芳烃 | Pseudoalteromonas(假交替单胞菌属)和 Agarivorans(食琼胶菌属)物种 | 这些菌株可能受碳氢化合物污染的影响而富集,并可能积极促进脂肪族碳氢化合物和多环芳烃的降解 | Dell'Anno et al. (2021) |
| 5 | 六价铬 | Escherichia coli(大肠杆菌) | 铬的降解 | Mohamed et al. (2020) |
| 6 | 黄曲霉毒素(一种剧毒真菌毒素) | P. aeruginosa(铜绿假单胞菌) | 黄曲霉毒素生物降解的潜力候选者 | Song et al. (2019) |
| 7 | 砷 | Microbacterium(微杆菌属)、Micrococcus(微球菌属)、Shinella(申氏菌属)和 Bacillus(芽孢杆菌属) | 金属生物修复 | Bhakat et al. (2019) |
| 8 | 铜、镉和铅 | Lysinibacillus(赖氨酸芽孢杆菌属)物种 | 降低了重金属浓度:铜降低 88%,镉降低 41%,铅降低 71% | Kharangate-Lad and D'Souza (2021) |
| 9 | 原油 | P. aeruginosa(铜绿假单胞菌) | 原油的生物修复 | Mukjang et al. (2022) |
| 10 | 重金属 | Saccharomyces cerevisiae(酿酒酵母)、Cunninghamella elegans(优美毛霉)、Rhodomicrobium(红微菌属)、Beggiatoa thiothrix(贝日阿托氏硫丝菌) | 重金属的生物修复 | Shah and Jain. (2020) |
| 11 | 氯乙烯 | Dehalococcoides(脱卤拟球菌属)物种 | 氯乙烯的生物修复 | Mohamed et al. (2020) |
| 12 | 多环芳烃 (PAHs) | Novosphingobium(新鞘氨醇单胞菌属)、Pseudomonas(假单胞菌属)、Comamonas(丛毛单胞菌属)、Burkholderia(伯克霍尔德菌属)、Bacillus(芽孢杆菌属)、Pseudoxanthomonas(假黄单胞菌属)、Paenibacillus(类芽孢杆菌属)和 Actinobacteria(放线菌门) | 多环芳烃的降解 | Mohapatra and Phale (2021) |
| 13 | 多溴二苯醚 (BDE) | P. chrysosporium(黄孢原毛平革菌) | P. chrysosporium 可能是去除 BDE 污染的理想微生物类型 | Liu et al. (2018) |
| 14 | 镉 | Cladosporium(枝孢菌属)、A. fumigatus(烟曲霉)、Paecilomyces(拟青霉属)、Terichoderma(木霉属)和 A. versicolor(杂色曲霉)物种 | 微生物对镉的潜在生物降解 | Unuofin et al. (2021) |
| 15 | 油脂 | Corynebacterium propinquum(邻近棒状杆菌)、P. aeruginosa(铜绿假单胞菌)、Alcaligenes odorans(嗅味产碱杆菌)和 Fusarium(镰刀菌属)物种 | 油脂的生物降解 | Pande et al. (2020) |
| 16 | 铬 | Ulothrix tenuissima(极细丝藻)、Spirulina(螺旋藻属)物种和 C. reinhardtii(莱茵衣藻) | 铬的生物降解 | Aregbesola et al. (2020) |
| 17 | 芳香烃 | Microbacterium(微杆菌属)、Pseudomonas(假单胞菌属)、Acinetobacter(不动杆菌属)和 Ralstonia(罗尔斯通菌属)物种 | 芳香烃的生物降解 | Basu et al. (2018) |
| 18 | 镉、铅和铬 | Aerococcus(气球菌属)物种和 Rhodopseudomonas palustris(沼泽红假单胞菌) | 镉、铅和铬的生物降解 | Sravya and Sangeetha (2022) |
| 19 | 苯酚 | B. Subtilis(枯草芽孢杆菌)、P. aeruginosa(铜绿假单胞菌)、Alcaligenes odorans(嗅味产碱杆菌)和 Corynebacterium propinquum(邻近棒状杆菌) | 苯酚的生物降解 | Gaur et al. (2018) |
| 20 | 还原染料 | K. oxytoca(产酸克雷伯菌)、B. firmus(坚强芽孢杆菌)、B. macerans(浸麻芽孢杆菌)和 Staphylococcus aureus(金黄色葡萄球菌) | 还原染料的生物降解 | Sangkharak et al. (2020) |
| 21 | 铅、镍和重铬酸盐 | Chlorella(小球藻属)和 Spirulina(螺旋藻属)物种 | 重铬酸盐、镍和铅的生物降解 | Geetha et al. (2021) |
| 22 | 重金属和汞 | Saccharomyces cerevisiae(酿酒酵母)和 Cunninghamella elegans(优美毛霉) | 重金属组分的生物降解 | Duc et al. (2021) |
| 23 | 染料 | Bacillus licheniformis(地衣芽孢杆菌) | 染料的生物降解 | Mousavi et al. (2021) |
| 24 | 铁和锌 | Pseudomonas fluorescence(荧光假单胞菌)和 Bacillus subtilis(枯草芽孢杆菌) | 铁和锌的生物降解 | Li et al. (2018) |
| 25 | 铬 | Bacillus(芽孢杆菌属)、Escherichia(埃希氏菌属)、Shewanella(希瓦氏菌属)、Enterobacter(肠杆菌属)、Thermus(嗜热菌属)和 Pseudomonas(假单胞菌属)物种 | 铬的生物降解 | Mousavi et al. (2021) |
| 真菌 (Fungi) | ||||
| 26 | 二甲苯、苯、甲苯、有机氯化合物和 N-杂环炸药 | Phanerochaete chrysosporium(黄孢原毛平革菌) | 有机化合物的生物降解 | Singh et al. (2020) |
| 27 | 4,4-二溴二苯醚 | Phanerochaete chrysosporium(黄孢原毛平革菌) | 4,4-二溴二苯醚的生物降解 | Sen et al. (2019) |
| 28 | 多环芳烃 (PAH) 和总石油烃 (TPH) | Aspergillus niger(黑曲霉)、Aspergillus flavus(黄曲霉)、Varicosporium elodeae(伊乐藻变异孢子菌)和 Cochliobolus lunatus(弯孢月黧菌) | 石油烃的降解 | Gaur et al. (2022) |
| 29 | 砷 | Saccharomyces cerevisiae(酿酒酵母) | 砷的生物降解 | Verma and Kuila (2019) |
| 30 | 砷 | Aspergillus(曲霉属)物种 | 砷的生物降解 | Mohd et al. (2019) |
| 31 | 4-羟基 -3,5-二氯联苯 | Coprinus comatus(毛头鬼伞) | 二氯联苯的生物降解 | Li et al. (2018) |
| 32 | 脂肪烃、多环芳烃和氯酚 | Aspergillus(曲霉属)和 Penicillium(青霉属)物种 | 脂肪烃、多环芳烃和氯酚的生物降解 | Li et al. (2020) |
| 33 | N-十六烷 | Aspergillus(曲霉属)物种 | N-十六烷的生物降解 | Al-Hawash et al. (2018) |
| 34 | 菲 | Phomopsis liquidambari(液巴拟茎点霉) | 菲的生物降解 | Fu et al. (2018) |
| 35 | 芘 | Ganoderma lucidum(灵芝) | 芘的生物降解 | Agrawal et al. (2018) |
| 36 | 钴和铜 | Trichoderma(木霉属)、Penicillium(青霉属)和 Aspergillus(曲霉属)物种 | 钴和铜的生物降解 | Dusenge et al. (2020) |
| 藻类 (Algae) | ||||
| 37 | 砷 | Microcystis aeruginosa(铜绿微囊藻) | 砷的生物降解 | Wang et al. (2018) |
| 38 | 砷 | Chlamydomonas reinhardtii(莱茵衣藻)和 S. almeriensis(阿尔梅里亚小球藻) | 砷的生物降解 | Saavedra et al. (2018) |
| 39 | 锌 | Fucus vesiculosus(墨角藻) | 锌的生物降解 | Brinza et al. (2020) |
| 40 | 铁 | Chlorococcum humicola(土壤绿球藻) | 铁的生物降解 | Chugh et al. (2022) |
| 41 | 苯酚 | Isochrysis galbana(等鞭金藻)、Phaeodactylum tricornutum(三角褐指藻)和 Chlorella(小球藻属)物种 | 苯酚的生物降解 | Wu et al. (2022) |
| 42 | 铬、镍、镉和铅 | F. vesiculosus(墨角藻) | 重金属的生物降解 | Moreira et al. (2019) |
| 43 | 镍和镉 | Cystoseria indica(印度囊藻) | 重金属的生物降解 | Moreira et al. (2019) |
| 44 | 铬和镉 | Chlamydomonas reinhardtii(莱茵衣藻) | 重金属的生物降解 | Nowicka et al. (2020) |
| 45 | 镉 | Microcystis aeruginosa(铜绿微囊藻) | 重金属的生物降解 | Deng et al. (2020) |
| 46 | 镉 | Cyclotella(小环藻属)、Scenedesmus protuberans(突起栅藻)和 Scenedesmus acuminatus(尖形栅藻) | 重金属的生物降解 | Vo et al. (2020) |
| 47 | 钴 | Chlorococcum humicola(土壤绿球藻) | 重金属的生物降解 | Chugh et al. (2022) |
四、基因工程微生物在生物修复中的作用
日益严峻的环境退化问题推动了生物修复技术及基因工程微生物(Genetically Engineered Microorganisms, GEMs)的快速发展。
与传统方法相比,利用工程微生物进行修复不仅成本更低,且安全性更高。通过基因编辑技术对细菌进行改造,使其表达具有特定强化功能的蛋白,从而构建出高效的 GEMs。
这些经生物技术改良的菌株能够迅速适应新型污染物或启动共代谢机制,其降解毒素的效率和速度往往显著优于天然菌株。
展望未来,基因工程技术在改善人类健康及修复生态环境方面展现出巨大的应用潜力。
五、生物修复的最新进展
5.1 基于组学(Omics)的生物修复工具
基因组学(Genomics)、转录组学(Transcriptomics)、蛋白质组学(Proteomics)和代谢组学(Metabolomics)等组学技术的深入理解与应用,极大地推动了生物修复领域的研究进程。
这些技术通过建立 DNA 序列与蛋白质、代谢物及 mRNA 表达量之间的关联,为评估原位生物修复过程提供了便捷而精准的手段(见图2)。
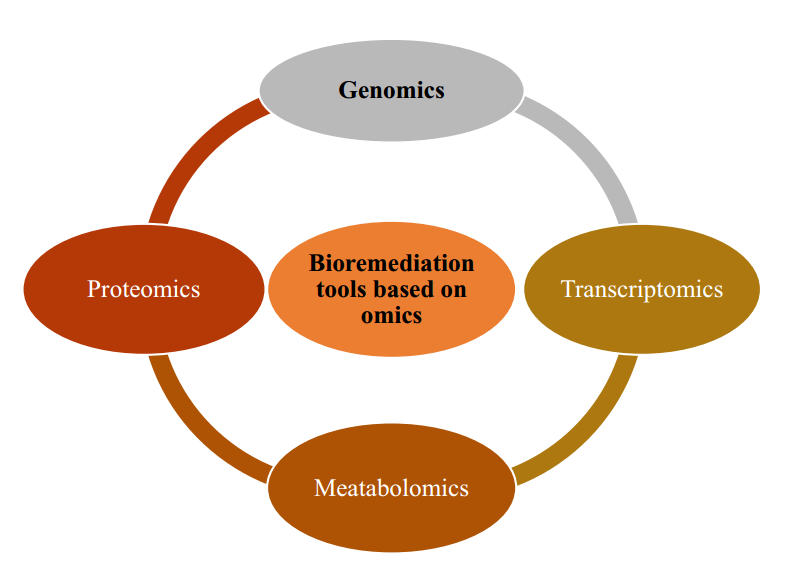
图2.基于组学的生物修复工具
5.1.1 基因组学 (Genomics)
基因组学方法有助于深入解析参与生物修复微生物的生理学特征。目前,大多数用于生物修复的微生物已拥有公开的全基因组或近乎完整的基因组序列。
利用全基因组 DNA 微阵列技术,研究人员不仅能识别这些完整序列,还能深入探究在污染物胁迫下的基因表达调控机制。
5.1.2 转录组学 (Transcriptomics)
转录组学利用微阵列分析等高通量技术,研究特定细胞或特定条件下由基因组转录生成的全部 RNA 集合(即转录组)。
通过转录组关联分析,可精准识别在不同微生物细胞中或针对不同处理条件表现出差异表达的基因。
作为连接基因组、蛋白质组与表型的桥梁,转录组学是分析 RNA 谱的有力工具。基因表达调控是微生物适应环境变化的核心机制。
DNA 微阵列技术的进步使得定量检测生物体内任意目标基因的 mRNA 表达水平成为可能。
然而,数据解读仍是微阵列实验面临的最大挑战:不同类型的胁迫通常会导致基因的上调或下调,这给统计分析带来了系统性误差和复杂性。
尽管如此,通过考察天然微生物群落的 mRNA 转录谱,转录组学及宏转录组学(Meta-transcriptomics)工具仍能为揭示微生物的活动机制提供宝贵见解。
5.1.3 代谢组学 (Metabolomics)
在生理代谢过程中,微生物会向周围环境释放特定的代谢产物。代谢组学旨在对这些化合物进行定性与定量分析。
利用核磁共振(NMR)和质谱(MS)光谱技术产生的特征峰图,可实现代谢物的精准检测;
此外,液相色谱和气相色谱等技术也广泛应用于此领域。代谢组学在以下方面具有重要应用价值:
- 揭示生态失衡机制:阐明污染物如何改变环境进而破坏宿主生态平衡;
- 解析信号传导: elucidate 细菌群体感应(Quorum sensing)期间的信号传递机制,以及微生物密度变化对基因表达的影响;
- 辅助药物研发:在代谢通路分析、新药发现及药物基因组学中发挥关键作用;
- 环境监测:作为评估环境健康状况的重要指标(Pulido et al., 2016)。
5.1.4 蛋白质组学 (Proteomics)
蛋白质组学通过研究生物体内的全套蛋白质,全面解析其在特定背景下的功能。当与其他组学技术联用时,它能提供对生物系统蛋白质组成的透彻理解,有助于揭示细胞过程的潜在机制,从而推动环境生物技术的发展。
与相对稳定的 DNA 不同,蛋白质组随时间及细胞状态动态变化,这使得蛋白质组学成为比基因组学更为精细和有力的研究工具。
通过对细菌培养物及环境样品中蛋白质序列的研究(即蛋白质组学和宏蛋白质组学 Metaproteomics),不仅能揭示蛋白质合成与组成的差异,还能发现多种对微生物应对污染物胁迫至关重要的功能蛋白。
特别是宏蛋白质组学,无需预先培养细菌即可直接测定环境样品中微生物群落的蛋白质表达谱,从而真实捕捉特定生态系统中微生物的实际功能活动。
随着二维凝胶电泳、蛋白质测序、质谱技术的进步以及蛋白质结构数据库的日益完善,该技术的应用价值愈发凸显。
六、结论
开发高效的生物修复策略依赖于对降解微生物群落的深入理解。多组学技术已成为发掘未知降解菌、解析代谢途径的关键工具。
研究表明,生物修复在效率、成本及生态友好性上均优于传统方法。未来需加强从实验室到大规模原位应用的转化研究,综合评估其经济与环境影响,以构建可持续的污染治理方案。
参考文献
1.Akinosho H, Yee K, Close D et al (2014) The emergence of Clostridium thermocellum as a high utility candidate for consolidated bioprocessing applications. Front Chem 2:66. https://doi.org/ 10.3389/fchem.2014.00066
2.Baughn AD, Malamy MH (2004) The strict anaerobe Bacteroides fragilis grows in and benefits from nanomolar concentrations of oxygen. Nature 427(6973):441–444.https://doi.org/10.1038/ nature02285
3.Chellapandi P, Bharathi M, Sangavai C et al (2018) Methanobacterium formicicum as a target rumen methanogen for the development of new methane mitigation interventions: a review. Vet Anim Sci 6:86–94. https://doi.org/10.1016/j.vas.2018.09.001
4.Krishna KV, Bharathi N, Malaviya A (2022) An updated review on advancement in fermentative production strategies for biobutanol using Clostridium species. Environ Sci Pollut Res 29: 47988–48019. https://doi.org/10.1007/s11356-022-20637-9
5.Najar IN, Sherpa MT, Das S et al (2020) Diversity analysis and metagenomic insights into the antibiotic resistance and metal resistances among Himalayan hot spring bacteriobiomeinsinuating inherent environmental baseline levels of antibiotic and metal tolerance. J Glob Antimicrob Res 21:342–352. https://doi.org/10.1016/j.jgar.2020.03.026
6.Verma G, Kumar V, Satyanarayana T (2022) Genomic attributes of thermophilic and hyperthermophilic bacteria and archaea. World J Microbiol Biotechnol 38:8.https://doi.org/10.1007/ s11274-022-03327-z
相关产品
HZB359393:地衣芽孢杆菌 | Bacillus licheniformis
HZB357530:恶臭假单胞菌 | Pseudomonas putida
HZB713192:铜绿假单胞菌 | Pseudomonas aeruginosa
HZB231920:酿酒酵母 CEN.PK2-1D | Saccharomyces cerevisiae CEN.PK2-1D
HZB119728:沼泽红假单胞菌 | Rhodopseudomonas palustris
HZB904321:产酸克雷伯菌 | Klebsiella oxytoca
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-03-12
编制人:思琪
审稿人:小藻