什么是原核生物?原核生物主要包括哪两大类群?
来源:武汉市灰藻生物科技有限公司 浏览量:5265 发布时间:2025-07-16 23:07:50
一、原核生物的特征
原核动物以单细胞或同源细胞群形式存在,尺寸通常为0.2-10微米,其分类依据是根据细胞构造与组分,而非生物体宏观属性。
除少数例外,其核质(基因带)与细胞质之间不存在核膜分隔。细胞分裂时,核质与细胞质不会发生周期性纹理或染色特性变化,也不形成微管纺锤体系统。质膜(细胞膜)常呈现复杂拓扑结构,会形成囊泡状、层状或管状结构内陷至细胞质中;独立于质膜系统的复制型细胞器(如绿硫菌囊泡、气胞)较为罕见,且由非单位膜包裹。
具备呼吸与光合功能的类群中,这些生理活动与质膜系统相关联,但蓝细菌的质膜与类囊体膜可能具有独立性。细胞质中散布70S型核糖体(古菌域核糖体沉降系数略高),不存在附着核糖体的内质网。细胞质无流动性,不表现胞质环流、伪足运动、内吞或外排现象。营养物质以分子形式获取。多数种类具有刚性细胞壁(非绝对特征)。运动方式包括无运动能力、游动(通过细菌型鞭毛驱动)或表面滑移运动。
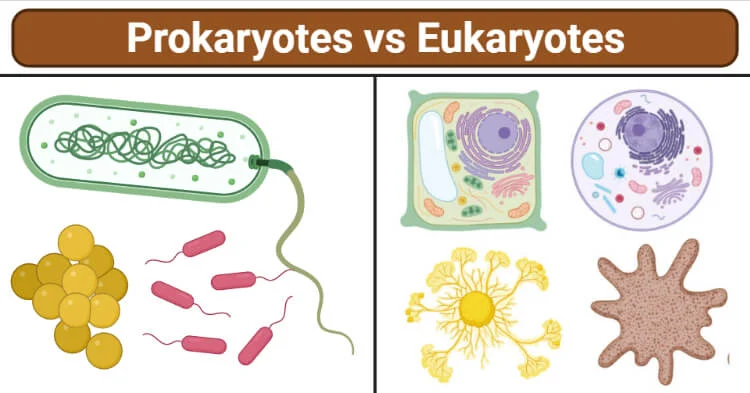
图1、原核生物和真核生物 来源: https://microbenotes.com/differences-between-prokaryotes-and-eukaryotes/
从生物体角度看,这些广泛分布于潮湿环境中的微生物主要为单细胞生物,但也存在丝状、菌丝体或群体形态。分化程度有限(可形成固着器、休眠细胞或特殊细胞形态)。存在基因转移与重组机制,但从不涉及配子形成与合子发育过程。
虽然原核生物通常易于与真核微生物区分,但在某些情况下可能难以辨别。尤其是当原核生物,表现出类似真核生物特征时。
例如:放线菌的菌丝,可能与霉菌菌丝混淆;细菌鞭毛束可能被误认为单根真核鞭毛;螺旋体的扭曲形态与某些原生动物的柔韧性相似;部分真核细胞尺寸与细菌相当,而某些细菌体型也可达到真核细胞水平(参见表1脚注)。最可靠的鉴别方法是证实核膜缺失(需透射电镜观察超薄切片),其他原核细胞特征从易检测指标,到需分子生物学手段验证的参数不一。
目前已开发出,可快速区分原核与真核细胞的荧光标记基因探针。
表1. 原核生物与真核生物的部分差异特征
| 特征 | 原核生物 | 真核生物 |
| 细胞学特征 | ||
| 核质(基因带、拟核)通过单位膜系统(核膜)与细胞质分离 | - | + |
| 细胞最小尺寸(宽度或直径): | ||
| 通常为0.2-2.0微米 | +b | - |
| 通常为2.0微米 | - | + |
| 存在线粒体 | - | 通常+ |
| 光养生物中存在叶绿体 | - | + |
| 若存在液泡,则由单位膜包裹 | - | + |
| 存在气泡c | D | - |
| 存在高尔基体 | - | D |
| 存在溶酶体 | - | D |
| 存在微管系统 | -d | D |
| 存在内质网 | - | + |
| 核糖体位置: | ||
| 分散在细胞质中 | + | - |
| 附着于内质网上 | - | + |
| 细胞质流动、伪足运动、内吞作用和外排作用 | - | D |
| 细胞分裂伴随核质或细胞质的纹理或染色特性周期性变化 | - | + |
| 鞭毛直径(若存在): | ||
| 0.01–0.02微米 | + | - |
| –0.2微米 | - | + |
| 横截面中,鞭毛具有“9+2”微管排列特征 | - | + |
| 存在内生孢子e | D | - |
| 对抗生素的敏感性 | ||
| 对青霉素、链霉素或其他原核生物特异性抗生素敏感 | D | - |
| 对环己酰亚胺或其他真核生物特异性抗生素敏感 | - | D |
| 基于化学分析的特征 | ||
| 存在聚β-羟基丁酸酯(作为细胞质内含物中的储存物质) | D | - |
| 存在磷壁酸(在细胞壁中) | D | - |
| 可能存在多不饱和脂肪酸(在膜中) | 罕见 | 常见 |
| 存在支链异或反异脂肪酸及环丙烷脂肪酸(在膜中) | 常见 | 罕见 |
| 存在甾醇(在膜中) | -f | 常见 |
| 特征 | 原核生物 | 真核生物 |
| 存在二氨基庚二酸(在细胞壁中) | Dg | - |
| 存在胞壁酸(在细胞壁中) | Dh | - |
| 细胞壁中存在肽聚糖(含胞壁酸) | Dh | - |
| 营养 | ||
| 细胞通过可溶性小分子获取营养;颗粒物或大分子需先被细胞膜外酶水解为小分子才能作为营养来源 | + | D |
| 代谢特征 | ||
| 呼吸和光合功能及相关色素和酶(如叶绿素、细胞色素)若存在,则与细胞膜或其内陷相关 | +i | - |
| 存在化能无机营养代谢(无机化合物可作为化学能生物的电子供体) | D | - |
| 具有固氮能力 | D | - |
| 能将NO3-异化为N2O或N2 | D | - |
| 产甲烷作用 | D | - |
| 能进行不产氧光合作用 | D | - |
| 酶学特征 | ||
| 超氧化物歧化酶类型 | ||
| Cu-Zn型 | 罕见 | + |
| Mn型和/或Fe型 | + | -j |
| 繁殖特征 | ||
| 细胞分裂包括有丝分裂,且存在微管(纺锤体)系统 | - | + |
| 减数分裂发生 | - | D |
| 基因转移和重组机制(若存在)涉及配子形成和合子形成 | - | + |
| 分子生物学特性 | ||
| 每个拟核/细胞核中的染色体数量 | 通常为1 | 通常为1 |
| 染色体呈环状 | + | - |
| 染色体呈线性 | -k | + |
| 核糖体沉降系数: | ||
| 70S | + | -l |
| 80S | - | |
| 核糖体RNA沉降系数: | ||
| 16S, 23S, 5S | + | - |
| 18S, 28S, 5.8S, 5S | - | + |
| 蛋白质合成中多肽链起始的第一个氨基酸: | ||
| 甲硫氨酸(Methionine) | D | + |
| N-甲酰甲硫氨酸(N-Formylmethionine) | D | - |
| 在16S或18S核糖体RNA的3'端存在AUCACCUCC结合位点 | + | - |
符号说明:
+ 表示存在或具有该特征;- 表示不存在或不具有该特征;D 表示因生物种类而异。
图2、原核生物 来源: https://microbenotes.com/differences-between-prokaryotes-and-eukaryotes/
二、古菌与细菌的差异
原核生物由两个根本不同的类群(域)组成:细菌(Bacteria)和古菌(Archaea)。
基于保守蛋白序列的系统发育分析表明,在大多数进化树中,古菌与真核生物(Eucarya)的亲缘关系,比古菌与细菌更近。
表2. 区分细菌与古菌的部分特征
| 特征 | 细菌 | 古菌 |
| 一般形态与代谢特征 | ||
| 严格厌氧菌,以 H2/CO2、甲酸盐、乙酸盐、甲醇、甲胺或 H2/甲醇为主要代谢底物生成甲烷。细胞在 420 nm 激发光下呈现蓝绿色荧光。 | — | D |
| 严格厌氧菌,通过异化硫酸盐还原生成H2S。极端嗜热(部分可在110°C下生长)。在420 nm激发光下呈现蓝绿色荧光 | — | D |
| 细胞呈革兰氏阴性或阳性,为好氧或兼性厌氧的化能有机营养体。形态包括杆状、规则或高度不规则。需高浓度NaCl(1.5 M及以上)。嗜中性或嗜碱性。中温或轻度嗜热(最高55°C)。部分物种含红紫色光活性色素菌视紫红质,可利用光生成质子动力 | — | D |
| 嗜热嗜酸、好氧、无细胞壁的球状细胞 | — | D |
| 专性嗜热、好氧、兼性厌氧或严格厌氧的革兰氏阴性杆状、丝状或球状菌。最适生长温度70°C至105°C。嗜酸或嗜中性。自养或异养。多数为硫代谢菌 | — | D |
| 细胞壁(若存在) | ||
| 含胞壁酸 | + | — |
| 抗生素敏感性 | ||
| 对β-内酰胺类抗生素敏感 | D | — |
| 脂类 | ||
| 膜磷脂组成:长链醇(植醇)通过醚键与甘油形成C20二植烷基甘油二醚或C40二双植烷基二甘油四醚 | — | + |
| 长链脂肪族脂肪酸通过酯键与甘油结合 | + | — |
| 脂质合成途径: | — | — |
| 甲羟戊酸途径 | + | — |
| 丙二酸途径 | 通常+ | — |
| 分子生物学特征 | ||
| tRNA“共同臂”含核糖胸腺嘧啶 | 通常 | + |
| tRNA“共同臂”含假尿苷或1-甲基假尿苷 | — | + |
| 蛋白质合成中多肽链起始氨基酸: | ||
| 甲硫氨酸 | — | + |
| N-甲酰甲硫氨酸(N-Formylmethionine) | + | — |
| 起始tRNA的氨基酸茎末端为“AU”碱基对 | — | + |
| 核糖体蛋白质合成受以下物质抑制: | ||
| 茴香霉素 | — | + |
| 卡那霉素 | + | — |
| 氯霉素 | + | — |
| 白喉毒素抑制肽链延伸因子EF-2的ADP核糖基化 | — | + |
| 延伸因子2(EF-2)含二吡啶酰胺氨基酸 | — | + |
| 部分tRNA基因含内含子 | — | + |
| DNA依赖性RNA聚合酶: | ||
| 多组分酶 | — | + |
| 受利福平及链霉溶菌素抑制 | + | — |
| 复制性DNA聚合酶受蚜栖菌素或丁苯基-dGTP抑制 | + | — |
2.1细菌
细菌域(Bacteria) 基于实用角度,细菌域可分为三种表型亚群:
(1)具有细胞壁的革兰氏阴性菌;
(2)具有细胞壁的革兰氏阳性菌;
(3)无细胞壁的菌种。
1.革兰氏阴性有壁细菌:
具有外膜与较薄肽聚糖层(含胞壁酸,少数退化类群例外)的细胞壁结构。形态包括球状、杆状、螺旋状或丝状,部分具有鞘膜或荚膜。繁殖以二分裂为主,少数出芽或多分裂(如蓝细菌II亚组)。运动方式包括游动、滑移或不运动。涵盖光能/化能自养与异养类型,包括需氧、厌氧、兼性厌氧及专性胞内寄生菌。
2.革兰氏阳性有壁细菌:
无外膜,肽聚糖层较厚。部分成员的细胞壁含有磷壁酸(teichoic acids)和/或中性多糖,少数类群(如含分枝菌酸的菌种)的细胞壁结构特殊。尽管通常经革兰氏染色呈阳性,但存在例外(如丁酸弧菌属因细胞壁异常薄而呈阴性反应)。细胞形态包括球形、杆状或丝状(杆状与丝状菌可呈非分枝或真分枝形态)。繁殖方式以二分裂为主,部分可形成休眠孢子(如内生孢子或菌丝孢子)。该大类涵盖非产孢菌、产孢菌、放线菌及其近缘类群。革兰氏阳性菌多为化能异养型,包括需氧、厌氧、兼性厌氧及微需氧种类,部分为专性细胞内寄生菌。唯一光合类群——螺旋杆菌虽具革兰氏阳性细胞壁但染色呈阴性。表3列出了区分革兰氏阳性菌与阴性菌的关键特征。
表3. 具有细胞壁的革兰氏阳性菌与具有细胞壁的革兰氏阴性菌的部分鉴别特征
| 特征 | 革兰氏阴性菌 | 革兰氏阳性菌 |
| 细胞学特征 | ||
| =存在外膜 | + | − |
| 抗酸染色 | − | Db |
| 存在内生孢子 | −c | Dd |
| 具有真分支的菌丝状生长 | − | D |
| 运动方式 | ||
| 滑行运动 | D | − |
| 化学特征 | ||
| 细胞壁干重中脂质占比 | 通常11-22% | 通常4%e |
| 存在磷壁酸或脂磷壁酸 | − | D |
| 细胞壁外膜中存在脂多糖(LPS)f | + | − |
| 存在2-酮基-3-脱氧辛酸(KDO)g | D | − |
| 细胞壁干重中肽聚糖占比 | 通常<10% | 通常10% |
| 存在分枝菌酸 | − | Dh |
| 存在磷脂酰肌醇甘露糖苷 | − | Di |
| 存在磷酸鞘脂 | Dj | − |
| 代谢特征 | ||
| 通过氧化无机铁、硫或氮化合物获取能量 | − | D |
| 酶学特征 | ||
| 柠檬酸合酶: | ||
| 受还原型烟酰胺腺嘌呤二核苷酸(NADH)抑制 | 通常+k | 通常− |
| 分子量: | ||
| ~250,000 | 通常− | 通常+ |
| ~100,000 | 通常+l | 通常− |
| 琥珀酸硫激酶分子量: | ||
| 70,000–75,000 | 通常− | 通常+ |
| 140,000–150,000 | 通常+ | 通常− |
注释:
a 本表列举了部分典型特征,实际可能存在例外。
b D表示因菌种而异(部分阳性菌为抗酸染色阳性,如分枝杆菌)。
c 革兰氏阴性菌通常不产内生孢子(例外:少数嗜热菌属)。
d 阳性菌中仅芽孢杆菌属和梭菌属等产孢子。
e 阳性菌细胞壁脂质含量较低,但含大量肽聚糖和磷壁酸。
f LPS为革兰氏阴性菌外膜特有成分,是内毒素主要来源。
g KDO常见于革兰氏阴性菌LPS核心多糖区。
h分枝杆菌等阳性菌含分枝菌酸。
i 放线菌等阳性菌含磷脂酰肌醇甘露糖苷。
j 拟杆菌等阴性菌含磷酸鞘脂。
k 阴性菌柠檬酸合酶通常受NADH抑制。
l 阴性菌柠檬酸合酶分子量通常为~100,000。
3.无细胞壁细菌(支原体)
它们不合成肽聚糖前体,且对β-内酰胺类抗生素,或其他抑制细胞壁合成的抗生素不敏感。其外层由单位膜(即质膜)包裹,细胞具有高度多形性,从可变形的大型囊泡到可通过滤器的微小(0.2微米)颗粒均有存在,分枝状丝状形态较为常见。繁殖方式可能包括出芽、断裂和/或二分裂。某些类群因内部结构排列而呈现一定规则形态。通常无运动能力,但部分物种具有滑行运动特性。目前尚未发现休眠体存在,革兰氏染色呈阴性。
大多数支原体需要复杂培养基(高渗透压环境)才能生长,在固体培养基表面,常形成特征性的"荷包蛋"状菌落。绝大多数还需胆固醇和长链脂肪酸,作为生长必需物质;若培养基中存在非酯化胆固醇,其会独特地整合进支原体膜结构(无论该菌株是否依赖固醇生长)。
其核糖体RNA的GC含量为43-48摩尔%(低于带细胞壁革兰氏阴性菌和阳性菌50-54摩尔%的水平);DNA的GC含量也相对较低(23-46摩尔%),基因组大小0.5-1.0×10^9道尔顿,小于其他原核生物。支原体可腐生、寄生或致病性生活,病原性支原体会引发动植物和组织培养物的疾病。
2.2 古菌
古菌的主要类群:
古菌主要为陆地及水生微生物,存在于厌氧、高盐、高温环境中;部分种类亦作为共生生物,存在于动物消化道。
古菌包含需氧菌、厌氧菌及兼性厌氧菌,可通过化能无机自养、有机营养或兼性有机营养方式生长。
其生长温度范围涵盖嗜温菌至嗜热菌,部分物种甚至可在110℃的高温环境中存活。
主要类群包括:
a) 产甲烷古菌
b) 硫酸盐还原古菌
c) 极端嗜盐古菌
d) 无壁古菌
e) 极端嗜热硫代谢古菌
所有古菌共有的独特生化特征是,其甘油类异戊二烯醚脂质结构。细胞壁中不含有胞壁质(含胞壁酸的肽聚糖),因此古菌对β-内酰胺类抗生素具有天然抗性。其tRNA的"共同臂"以假尿苷或1-甲基假尿苷替代了核糖胸苷。5S、16S和23S rRNA的核苷酸序列与细菌域、真核生物域存在显著差异。
古菌的独有生化特征包括:
1.含甘油异戊二烯醚脂质
2.细胞壁无胞壁酸(对β-内酰胺抗生素不敏感)
3.tRNA"共同臂"含假尿苷/1-甲基假尿苷
4.rRNA序列与细菌/真核生物差异显著
古菌与真核生物的共有分子特征包括:
(a) 延伸因子2(EF-2)含有二吡啶酰胺氨基酸,因此可被白喉毒素ADP-核糖基化;
(b) 核糖体"A"蛋白的氨基酸序列与真核生物对应蛋白(L-7/L12)具有同源性;
(c) 甲硫氨酸起始tRNA未被甲酰化;
(d) 部分tRNA基因含有内含子;
(e) 起始tRNA的氨酰茎末端以"AU"碱基对结尾;
(f) DNA依赖性RNA聚合酶为多组分酶,且对抗生素利福平及链霉溶菌素不敏感;
(g) 与真核生物的α-DNA聚合酶类似,古菌的复制DNA聚合酶不受蚜栖菌素或丁苯基-dGTP抑制;
(h) 蛋白质合成受茴香霉素抑制,但不受氯霉素影响。
自养型古菌并不通过卡尔文循环同化CO₂。在产甲烷菌中,自养CO₂通过一条涉及独特辅酶的途径被固定,这些辅酶包括甲烷呋喃、四氢甲烷蝶呤、辅酶F420、HS-HTP、辅酶M、HTP-SH和辅酶F430;而在酸菌属(Acidianus)和热变形菌属(Thermoproteus)中,自养CO₂则通过还原性三羧酸途径固定。部分产甲烷古菌具有固氮能力。
革兰氏染色结果在同一亚群中可能呈现阳性或阴性,这是由于细胞包膜类型差异显著所致。革兰氏阳性染色物种具有假胞壁质、甲烷软骨素和杂多糖细胞壁;革兰氏阴性染色细胞则具有(糖)蛋白表面层。其细胞形态多样,包括球形、螺旋形、板状或杆状;单细胞和多细胞形式(如丝状或聚集体)亦存在。单个细胞直径范围为0.1-15微米,丝状体长度可达200微米。繁殖方式包括二分裂、出芽、缢缩、断裂或未知机制。菌体颜色可呈现红色、紫色、粉色、橙棕色、黄色、绿色、绿黑色、灰色及白色。
参考资料
Bergeys Manual of Systematic Bacteriology (Garrity, Berner, Creig) . Volume Two:The ProteobacteriaPart A Introductory Essays.
Bergey’ s Manual of Determinative Bacteriology.
敬请关注“灰藻视界”,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2025-07-15
编制人:思琪 | 审稿人:小藻