蓝细菌、立克次体、支原体、衣原体、螺旋体、蛭弧菌及黏细菌介绍
来源:武汉市灰藻生物科技有限公司 浏览量:10542 发布时间:2025-04-28 13:55:23
蓝细菌
蓝细菌曾称蓝藻、蓝绿藻,是一类进化历史悠久、革兰氏阴性、无鞭毛、含叶绿素 a(不形成叶绿体)、具胡萝卜素及藻胆蛋白,且能进行产氧性光合作用的大型原核微生物。蓝细菌营养简单,分布极广,在河流、海洋、湖泊和土壤中生长,在极端环境(如温泉、盐湖、荒漠、岩石表面或风化壳中及植物树干等)中也能生长,称为“先锋生物”。
蓝细菌在地球上已存在了约 35 亿年,可能是地球上最早的产氧的光合生物,对地球大气从无氧转变为有氧及在真核生物的进化等过程中起重要作用。蓝细菌与人类关系密切,有重要的经济价值,近年来研究较多的螺旋藻是由钝顶螺旋蓝细菌(Spirulina platensis)和极大螺旋蓝细菌(S. maxima)等加工成的一种营养食物,富含蛋白质、多糖、维生素、钙、铁、微量元素、β-类胡萝卜素及 y-亚麻酸等,对肝硬化、贫血、白内障、青光眼、胰腺炎、糖尿病、肝炎等疾病有一定的辅助治疗作用。普通木耳念珠蓝细菌(Nostoccommune)(地木耳)和发菜念珠蓝细菌(N. flagelliforme)等都可食用。有些蓝细菌还能与真菌、苔藓、蕨类、珊瑚和种子植物共生,如地衣就是蓝细菌与真菌的共生体,红萍是固氮鱼腥蓝细菌(Anabaena azotica)和蕨类植物满江红(Azolla imbricata)的共生体。已知固氮蓝细菌有 120 多种,在岩石风化、土壤形成及增加土壤氮营养、沙漠化土壤治理、促进植物生长等方面有重要作用。蓝细菌能结合并清除水中的有害金属和化学物质,淡化海水,监测环境污染,处理污水等。
蓝细菌形态多样,有的是二分裂或复分裂形成的,蓝细菌的形态和大小球状或杆状的单细胞,有的呈丝状体。其细胞比细菌大,直径3.0~10.0μm,巨颤蓝细菌(Oscillatoria princeps)直径达60.0μm。
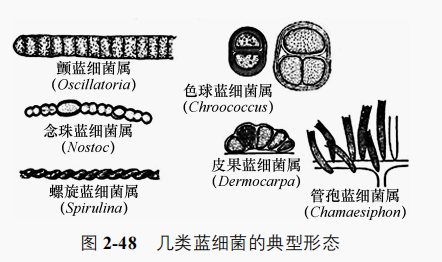
蓝细菌细胞壁有内外两层,外层为脂多糖层,内层为肽聚糖层。许多蓝细菌还向细胞壁外分泌黏胶物质,形成类似细菌荚膜的外套或包裹在丝状体外形成鞘。多数丝状蓝细菌虽无鞭毛,但能做滑行运动。细胞膜单层,很少有间体。其脂肪酸一般是含有两个或多个双键的不饱和脂肪酸,其他细菌通常只含有饱和脂肪酸及只有一个双键的不饱和脂肪酸。细胞核无核膜,核糖体 70S。
蓝细菌的营养要求简单,不需要维生素,能以硝酸盐或氨化物作氮源,有些种能固定空气中的氮。其光合作用部分称类囊体,数量很多,以平行或卷曲的方式贴近细胞膜。类囊体膜上有叶绿素a及β-胡萝卜素、藻胆素等辅助光合色素和电子传递链的有关组分。藻胆素为蓝细菌所特有,缺氮时可作氮源。藻胆素与蛋白质共价结合成藻胆蛋白(PBP),聚集在类体外表面构成颗粒状藻胆蛋白体。胆蛋白含藻蓝素和藻红素两种色素,不同的蓝细菌各种色素比例不同,故呈现绿、蓝、红等不同的颜色。大多数蓝细菌兼有藻蓝素和叶绿素 a,使细胞呈特殊的蓝色,故称蓝细菌。但受氮饥饿的蓝细菌,由于藻蓝素被用去,所以常呈绿色。有些蓝细菌呈红色或棕色系藻红素所致。蓝细菌的藻蓝素和藻红素比例受生长环境的影响,尤其是光照的变化。在蓝、绿光下,藻红素占优势,在红光下主要是藻蓝素,这就保证了蓝细菌对不同生境的适应性。有些蓝细菌的代谢产物对环境有重要影响,如许多蓝细菌能产生神经毒素,动物饮用了含大量这类蓝细菌的水后会很快死亡。蓝细菌细胞内有各种贮藏物,如糖原、聚磷酸盐、PHB、氮源储藏物--蓝细菌肽,还有能固定CO2的羧酶体。水生性种类细胞中常有气泡,能保持细胞浮在上层水面,得到最充足的光线,以利光合作用。
蓝细菌细胞特化形式
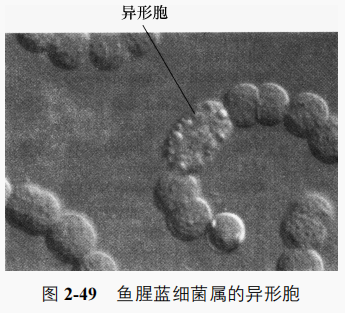
(1)异形胞分布在丝状体中间或末端,比营养细胞大、色浅,细胞壁变厚(图 2-49),环境中缺乏硝酸盐或氨时由少数营养细胞异化而来,是适应在有氧条件下固氮的细胞。它仅含少量的藻胆蛋白,只存在光合系统I,缺少产氧的光合系统Ⅱ,不会因光合作用而产生对固氮酶有严重毒害的分子氧,却能产生对固氨所必需的 ATP。异形胞与邻近的营养细胞间由厚壁孔道相连,利于“光合细胞”和“固氮细胞”间的物质交流。有些不形成异形胞的藻类如能产生鞘膜的黏球蓝细菌属中的几种单胞藻在有氧条件下也可以固氮,说明在其细胞中有一套有效的除氧系统以保证固氨酶的活力。
(2)静息孢子是一种在干燥、低温和长期黑暗等条件下长在细胞链中间或末端的形大、色深、壁厚的休眠细胞,富含贮藏物,能抵抗冷冻和干旱等不良条件。
(3)链丝段 又称藻殖段,是长细胞链断裂而成的短链段,有繁殖功能,可形成新的菌丝。
(4)内孢子少数种类[如管孢蓝细菌属(Chamaesiphon)]能在细胞内经复分裂形成许多球形或三角形的内孢子,待成熟后即可释放,具有繁殖功能,可形成新的营养细胞。
3.蓝细菌的繁殖
蓝细菌以无性方式繁殖。单细胞类群以裂殖(二分裂或多分裂)方式繁殖;有异形胞的丝状蓝细菌形成静息孢子;丝状类群除能通过裂殖使丝状体加长外,还能通过含有两个或多个细胞的链丝段脱离母体后长成新的丝状体。少数类群可通过形成内孢子繁殖。
4.蓝细菌的分群蓝细菌根据形态特征可分为5群。前两群为单细胞或呈团聚体,后三群呈丝状聚合体。
(1)色球蓝细菌群细胞呈球状或杆状,单生或聚合体;细胞间有英膜或黏液;以裂殖或出芽繁殖。代表有聚球蓝细菌属(Sjnechococcus)、黏球蓝细菌属(Gloeocapsa)等。
(2)宽球蓝细菌群包括仅通过复分裂繁殖的单细胞蓝细菌。在鞘套内排成丝状的杆状单细胞借复分裂产生许多小球状有繁殖能力的细胞。该群的代表有宽球蓝细菌属(Pleurocapsa)、皮果蓝细菌属(Dermocarpa)等。
(3)颤蓝细菌群细胞链仅由营养细胞组成,在丝状鞘套内排成链状的球状单细胞借二分裂和菌丝断裂繁殖,如螺旋蓝细菌属(Spirulina)、鞘丝蓝细菌属(Lvmngbya)等。
(4)念珠蓝细菌群是唯一能进行二分裂并产生异形胞的丝状蓝细菌。细胞链在只有游离氮作氮源时会分化出异形胞,有时产生静息孢子,如鱼蓝细菌属(Anabaena)等。
(5)分支异形孢蓝细菌群是能多平面多方向分裂、有异形胞的丝状蓝细菌。各属都能产生分支,有些属形成多个细胞列。有些产生静息孢子,如费氏蓝细菌属(Fischerella)。
立克次体
立克次体是一类介于细菌与病毒之间、接近于细菌、专性真核活细胞内寄生的原核微生物。它与支原体的区别是有细胞壁和不能独立生活。与衣原体的区别是其细胞较大、无过滤性和有不完整的产能代谢系统,不形成包涵体,必须通过媒介感染宿主。
立克次体一般呈球状或杆状,球状直径0.2~0.5μm,杆状大小为(0.3~0.5)μmx(0.8~2.0)μm。在不同宿主中或不同发育阶段可表现出不同形态,如球状、双球状、杆状或丝状等(图2-50)。不运动,无芽孢,革兰氏染色阴性,光学显微镜下可见,不同的种寄生在细胞不同部位,常存在于宿主细胞质或细胞核中。立克次体基因组很小,如普氏立克次体(Rickettsia prowazekii)基因组为 1.1Mb,含834个基因。其细胞结构及化学组成与G-菌相似,以二分裂方式繁殖,繁殖速度慢。
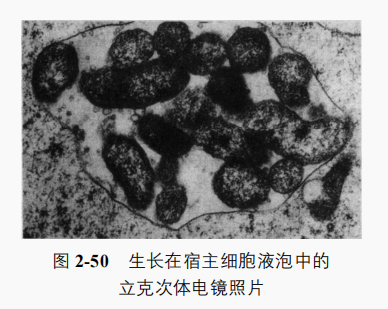
立克次体缺少糖酵解基因及合成氨基酸和核苷酸的许多基因,仅能进行一部分独立的代谢活动,如可以氧化谷氨酸;有细胞色素系统,可进行氧化磷酸化反应。酶系统不完全,如缺少糖酵解途径,不能氧化葡萄糖、6-磷酸葡萄糖、有机酸,故不能脱离宿主细胞独立生活另外,立克次体的细胞质膜比较疏松,物质较易通过,这既对它吸取宿主的大分子营养物质有利,也使其细胞物质易外渗,一旦脱离宿主会立即死亡。因此,除战热立克次体(Rquintana)可在没有活细胞的培养基中培养外,其他立克次体均不能在人工培养基上生长,必须在活细胞内才能生长繁殖,通常在敏感动物、鸡胚及动物组织中培养。1934年,我国科学家谢少文首先用鸡胚成功培养立克次体,为认识和防治立克次体病作出了重大贡献。
立克次体对热、光照、干燥及化学药剂抗性差,60℃处理 30min 即死亡,100℃很快死亡对消毒剂、磺胺及多种抗生素敏感,四环素、氯霉素、红霉素等可抑制其生长。
立克次体常寄生在虱、蚤、蜱、螨等节肢动物的消化道上皮细胞内,但不会致病。人的立克次体要通过这些动物叮咬、抓伤或随昆虫的粪便或口器进入血液而传播,在血细胞中大量繁殖可导致细胞破裂,并产生内毒素。其内毒素耐低温、干燥,但对热、消毒剂敏感。因此,防治人立克次体病的主要措施是消灭虱、蚤等传染媒介。
人立克次体病主要有流行性斑疹伤寒、落基山斑疹伤寒、恙虫热和Q热等。引起流行性斑疹伤寒的病原体是普氏立克次体,由虱传播;引起落基山斑疹伤寒的立氏立克次体(R.rickettsii)由蜱传播;引起人恙虫热的恙虫热立克次体(R tsutsugamushi)由螨传播;引起地方性斑疹伤寒的穆氏立克次体(R. mooseri)由蚤传播;Q热的病原体是伯氏考克斯体(Coxiella burneti)立克次体中抗性较强,在宿主体外也能存活。立克次体也寄生在植物细胞中,称类立克次体。
支原体
支原体是介于细菌与立克次体之间、无细胞壁的原核微生物。最早从患胸膜肺炎的牛体内分离,它们在液体培养基中培养常呈分支的丝状体,故称支原体。它是已知的能离开活细胞独立生活的最小生物,是最小的原核微生物,大小只有 0.10~0.25μm。其基因组很小,仅为0.6~1.1Mb。突出的结构特征是没有细胞壁,有细胞膜。细胞柔软,形态多变,呈球状或长短不一的丝状及分支状,对渗透压敏感。能通过细菌过滤器。其细胞膜为三层单位膜结构,内外两层均为蛋白质和糖类,中层为类脂、胆固醇和类胡萝卜素。质膜中有较多的甾醇或脂多糖,比其他原核生物的膜更坚韧。
支原体革兰氏染色阴性;不产生芽孢;无鞭毛;大多数不能运动,但能沿液体表面滑行;以二分裂方式繁殖,有些可以出芽的方式繁殖或从球状体长出丝状体,丝状体断裂成球状进行生长循环。多数支原体营兼性厌氧生活,多数种类能以糖作碳源。固体培养基上生长大多数支原体的菌落呈典型的“油煎蛋”状,中央厚实,颜色较深,这是由于中央菌体向培养基深部生长,四周扁平透明。菌落很小,在液体培养基中生长不产生混浊。人工培养需要较丰富的营养,通常要加入新鲜血清及牛心、酵母浸出汁,提供固醇等。支原体对热及干燥抵抗力弱,45℃处理30min 即可被杀死。对苯酚、来苏尔等化学消毒剂及各种表面活性剂、脂溶剂等敏感。支原体的生长不受青霉素、环丝氨酸等抑制细胞壁合成的抗生素影响,对溶菌酶不敏感。但对四环素等抑制蛋白质合成的抗生素和两性霉素等破坏含甾醇的细胞膜的抗生素都敏感。
据对固醇的需要将支原体分两类:一类为需固醇类群,包括支原体属(Mycoplasma)、厌氧支原体属(Anaeroplasma)、螺原体属(Spiroplasma)和脲原体属(Ureaplasma);另一类为不需固醇类群,有无胆甾原体属(Acholeplasma)和热原体属(Thermoplasma)。
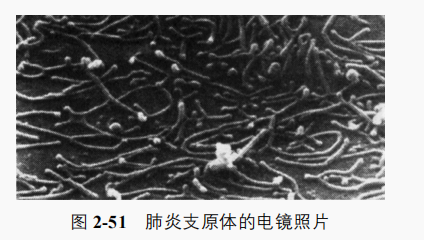
支原体广泛分布于动植物体内外及土壤、污水中,有寄生的,有腐生的。少数是动植物的病原菌,如丝状支原体丝状亚种 SC型(Mycoplasma mycoidessubsp.mycoides SC)是牛传染性胸膜肺炎病原体;无乳支原体(M agalactiae)引起羊的缺乳症;肺炎支原体(M.pneumoniae)引起人非典型肺炎,其主要通过飞沫传播;人型支原体(M.hominis)引起女性泌尿及生殖道感染。近来发现有些支原体是植物病害病原体,称类支原体或植原体,高等植物感染后的症状主要是丛枝(如泡桐丛枝病)、黄化(如翠菊黄化病、水稻黄萎病)和组织坏死(如橡胶褐皮病)等。
支原体与L型细菌相似,鉴定支原体前应在无抗生素培养基连续转接5次以排除后者。
衣原体
衣原体是介于立克次体与病毒之间的一类原核微生物,能通过细菌过滤器,在真核细胞内专性寄生。以前一直把这类微生物误认为是大型病毒,直至 1956 年我国著名微生物学家汤飞凡等通过鸡胚卵黄囊接种培养,在世界上首先成功从沙眼中分离出该病原菌并对它进行研究,逐步证实它是一类独特的接近于细菌的原核生物。衣原体已可用多种细胞培养。
衣原体的形态与立克次体相似,球形或椭圆形;较立克次体稍小,直径为0.2~0.3μm,在光学显微镜下勉强可见;细胞壁主要由脂多糖和蛋白质组成,缺少肽聚糖层;同时含有 DNA和 RNA,基因组分子量大小为(4~6)x108:核糖体为70S;以二分裂繁殖。
衣原体有一定的独立代谢能力,但生物合成能力差,更突出的是它们缺乏 ATP 再生体系,不能磷酸化葡萄糖或代谢葡萄糖,它们能吸收宿主细胞的 ATP 和 CoA。因此,衣原体必须在活细胞中生长繁殖,从宿主细胞获得能量才能生活,故又称“能量寄生物”
衣原体生活周期包括大小不同的两种细胞类型。小细胞称原体,球状,直径小于 0.4μm,壁厚、致密、不能运动、不生长(RNA:DNA=1∶1),能在宿主细胞外存活,抗干旱,有传染性,通过胞饮作用进入宿主细胞,被宿主细胞膜包围形成空泡,原体逐渐长大为始体。大细胞称始体或网状体,球状,直径1.0~1.5μm,壁薄而脆弱,易变形,G-菌,无传染性。利用宿主的高能化合物和小分子中间代谢物合成自己的细胞物质,生长较快(RNA:DNA-3∶1),通过二分裂反复繁殖形成大量子细胞。子细胞又变成原体聚集于细胞质内成为各种形状的包涵体,宿主细胞破裂后释放,再感染新的宿主细胞(图2-52)。整个周期约 48h。
![]()
衣原体抗性差,对热敏感,56~60℃存活5~10min。冰冻条件下可存活数年。对消毒剂和四环素、氯霉素、红霉素、多西环素及磺胺等敏感。鹦鹉热衣原体(Chilamydia psittaci)对磺胺有抗性。衣原体可直接侵入宿主细胞,能感染人类、鸟类及哺乳动物。例如,沙眼衣原体(C.trachomatis)为人类沙眼、埃及眼疾(结膜炎并导致失明)的病原体:肺炎衣原体(Cpneumoniae)是非典型肺炎的病原体,可引起肺炎并严重损伤心脏;鹦鹉热衣原体是鸟类肠胃道及呼吸道感染的病原体,可从病鸟传染给人,使人患肺炎并侵染消化道、生殖道。
立克次体、支原体、衣原体与细菌和病毒特性的比较见表 2-5。
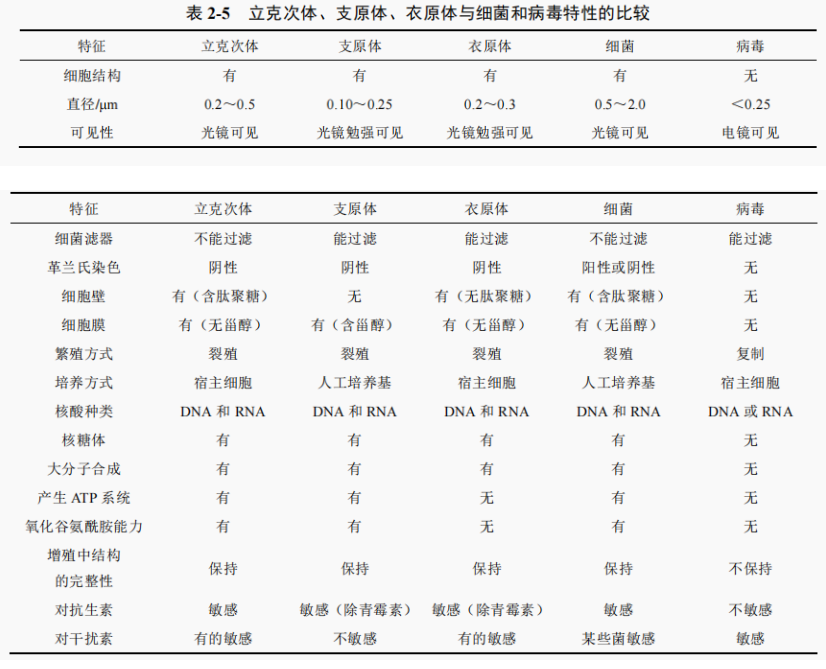
螺旋体
螺旋体是一群菌体细长并弯曲成螺旋状、运动活泼、介于细菌和原生动物之间的单细胞原核微生物。细胞非常长[(0.1~3.0)μmx(3.0~500.0)μm],有细胞壁但不及细菌坚韧,菌体柔软,不产生芽孢,革兰氏染色阴性。除个别种类如钩端螺旋体属(Leptospira)好氧外,其余均为厌氧或兼性厌氧型。能在含糖、蛋白胨、酵母膏、还原剂和脂肪酸的培养基上生长。有的不能在人工培养基上生长。
螺旋体具备细菌所有的基本结构,主要由原生质柱、轴丝和外鞘组成。外包细胞膜与细胞壁的原生质柱螺旋状,是螺旋体细胞的主要部分,内含细胞质和拟核。轴丝着生于原生质柱两个亚极端的细胞膜,缠绕在原生质柱外向另一端延伸,在其中部重叠,外包有外鞘。外鞘只能在超薄切片的电镜照片或负染色标本中观察到,是由蛋白质、脂类和碳氢化合物组成的柔软多层膜。外鞘破损菌体就死亡。轴丝数一般2~100条。轴丝的超微结构(基部有“钩”,有成对的盆状结构)、化学成分(亚基螺旋排成的蛋白质)和着生方式(一端连于细胞,一端游离)均与鞭毛相似,称周质鞭毛。螺旋体借轴丝快速旋转使菌体表面的螺旋凸纹不断伸缩而移动,由此推动细胞呈拔塞钻状快速前进。若是游离细胞外鞘沿纵轴旋转,细胞向前运动:如固着在固体表面,外鞘转动,细胞就向前爬行。
螺旋体以二分裂方式繁殖。螺旋体广泛分布于水体和动物体内,可分腐生和寄生两类,大多数属腐生类群,生活在水体、污泥和垃圾中。寄生类群有致病和非致病两类。哺乳动物肠道、睫毛表面、白蚁和石斑鱼肠道、软体动物躯体及反刍动物瘤胃中都有螺旋体。有些是动物体内固有的微生物区系,有些有致病性,如梅毒密螺旋体引起梅毒,回归热螺旋体引起回归热,伯氏疏螺旋体引起慢性游走性红斑(莱姆病),钩端螺旋体引起人的钩端螺旋体病(图 2-53)。根据形态、生理特性、致病性和生态环境,螺旋体分为 13 个属。其中重要的属为螺旋体属(Spirochaeta)、脊螺旋体属(Cristispira)、密螺旋体属(Treponema)、疏螺旋体属(Borrelia)、钩端螺旋体属(Leptospira)、纤细螺旋菌属(Leptonema)短螺旋体属(Brachyspira)和蛇形螺旋体属(Serpulina)等。螺旋体在分类上列为螺旋体门。
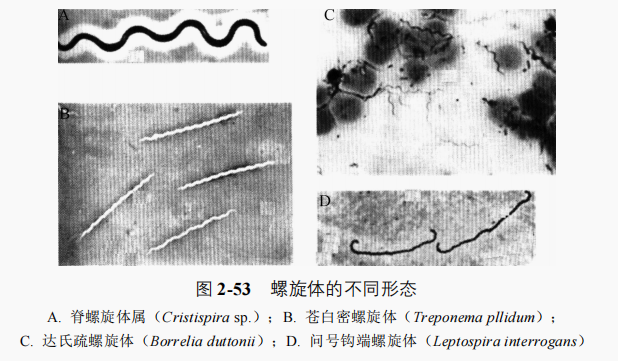
蛭弧菌
蛭弧菌是个体微小、可寄生并裂解其他菌的特殊原核微生物。基本特征与细菌相似,单细胞,弧形或逗号状,G-菌,专性好氧,(0.3~0.6)μmx(0.8~1.2)μm,能通过细菌过滤器并形成蛭弧菌斑。多为端生单鞭毛,水生类群的鞭毛外还附有由壁延伸形成的鞘膜,运动活跃。它借特殊的“钻孔”效应(还有酶的作用)进入后杀死宿主细菌,利用宿主的细胞质为营养,在周质空间生长为螺旋状蛭弧体,宿主细胞膨大呈球状,蛭弧体最后均分为多个具鞭毛的子细胞随宿主细胞裂解而释放,通常释放5~6个蛭弧菌。完成全过程需 2.5~4.0h
在固体培养基平板上,蛭弧菌也能形成类似噬菌斑的斑点,不过噬菌斑只有在宿主细胞生长时才会变大,而蛭弧菌斑能在被侵袭细胞停止生长后仍继续增大,最终在平板表面形成一个大斑,可呈现一定的颜色。可以从这一蚀斑中分离到蛭弧菌的纯培养。可用添加高浓度细菌提取物的无敏感细菌的培养基培养蛭弧菌。
蛭弧菌可侵袭 G-菌,不侵袭G+菌。部分类群也可腐生,不能利用葡萄糖产能,以蛋白质.肽、氨基酸和乙酸钠为能源和碳源,能在含酵母膏和蛋白胨的天然培养基上生长。专性好氧,适宜在 pH 6.0~8.5、温度 23~37℃的环境中生活。蛭弧菌广泛存在于土壤、河流、污水及近海水域中,它们在污染环境的净化和动植物细菌性病害防治等方面具有一定的应用价值。
黏细菌
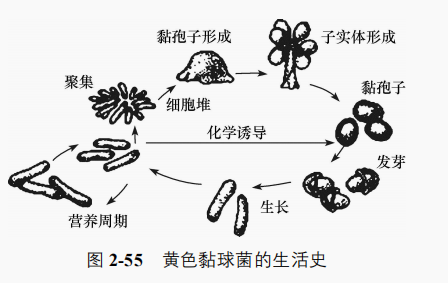
黏细菌是细菌中具有复杂的行为模式和生活周期的类群。典型黏细菌的生活周期可分为营养细胞和子实体两个阶段。营养细胞杆状,宽小于 1.5μm,G-菌,突出的特点是在体外产生黏液物质并将细胞团包埋在黏液中,无鞭毛,能借黏液在固体或气液界面上滑行。营养细胞以二分裂繁殖,缺乏营养时能趋聚成团,形成形态各异、颜色鲜艳、肉眼可见的子实体。营养细胞在子实体内转变为球形的黏孢子,有较强的抗性和折光性,并能在子实体干后借风和水等传播,在适宜的条件下萌发为营养细胞。
黏细菌专性好氧,主要以死的或活的细菌、真菌和藻类细胞或有机物为养料,多数能在含蛋白胨或酪蛋白水解物的固体培养基上生长。产生的胞外酶能水解蛋白质、核酸、脂肪和各种糖,有的能分解纤维素。多数种能溶解真核和原核生物。许多黏细菌可分泌抗生素杀死捕食者。主要分布在土壤表层、树皮、腐烂木材、堆厩肥和食草动物粪便上。
黏细菌在原核生物中生活周期和群体变化最复杂,对研究微生物的进化发育等有重要价值;能产生丰富的次级代谢产物,能分解复杂有机质。其代谢产物结构新颖,作用机制独特。纤维堆囊菌(Sorangium cellulosum)产生的埃博霉素能抗肿瘤,水溶性好,副作用小。近十多年从黏细菌中发现 600 余种具不同生理功能的活性物质,已成研究热点,开发应用潜力巨大。
参考资料
蔡信之, 黄君红, 康贻军. 《微生物学(第四版)》. 北京: 科学出版社, 2023.
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2025-04-28
#创作团队
编制人:木木 | 审稿人:小藻