古菌的形态、结构、遗传学特征及类群
来源:武汉市灰藻生物科技有限公司 浏览量:4261 发布时间:2025-03-03 15:33:12
引言
20 世纪 70 年代末,Woese 等在利用 16SrRNA 等序列研究原核生物间的相互关系时,发现它们中间存在差异,就提出将原核生物的另一部分称为古细菌,又简称为古菌。古菌多数生活在高温、低温、高酸、高碱、高盐、高压及高辐射等恶劣环境中,它们不仅能耐受这些极端环境,而且能在这类特殊生境中生长繁殖,甚至为了更好地繁衍后代需要一种或多种极端条件。少数种可作为共生菌存在于动物消化道内。
极端环境下微生物的生态、结构、分类、代谢、遗传等与一般环境中的微生物都有区别,研究极端环境微生物有重要的学术价值,特别是对研究在地球早期恶劣自然环境中生命起源和生物进化有重要意义。记载的古菌已有 289种(2006年)。我国的极端环境微生物资源丰富,开发利用潜力巨大。
一、古菌的形态和大小
古菌的形态多样,有球形、杆形、螺旋形、方形、三角形、棒状、盘状和叶状等。
古菌直径在 0.1~15.0um,长度可达 200um,有的需用电子显微镜才能观察清楚。
二、古菌的细胞结构
1.细胞壁:
古菌细胞壁的功能与真菌类似,其组成比较多样,不含胞壁酸、D型氨基酸和二氨基庚二酸。G-古菌细胞壁由蛋白质或糖蛋白亚基组成,没有外壁层和肽聚糖网状结构。G+古菌细胞壁与 G+菌相似,但古菌细胞壁由多糖(假肽聚糖)、糖蛋白或蛋白质构成,无真正的肽聚糖。古菌对溶菌酶及抑制肽聚糖合成的青霉素等有抗性。
(1)假肽聚糖细胞壁其结构与肽聚糖相似,多糖骨架由Ⅳ-乙酰葡糖胺和 乙酰塔罗糖胺糖醛酸以 B-1.3-糖苷键交替连接而成,肽尾由 L-Glu、L-Ala和 L-Lys 三个氨基酸组成,肽桥由一个 -Glu 组成,如甲烷杆菌属(Methanobacterium)(图 2-46)
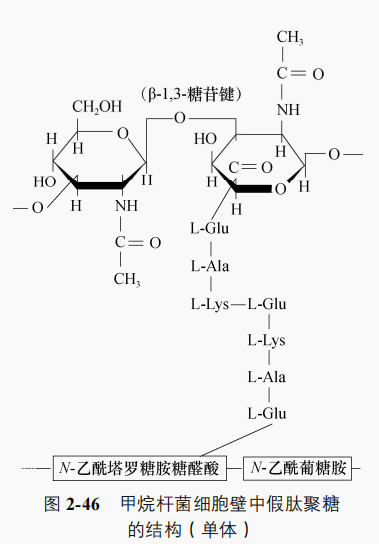
(2)独特多糖细胞壁其中含有独特的多糖,该糖含半乳糖胺、葡萄糖醛酸、葡萄糖和乙酸,不含磷酸和硫酸。例如,甲烷八叠球菌属(Metha-nosarcina)细胞壁,革兰氏反应阳性。
(3)硫酸化多糖细胞壁极端嗜盐古菌盐球菌属(Halococcus)的细胞壁是由硫酸化多糖组成的。其中含葡萄糖、甘露糖、半乳糖和它们的氨基糖,以及糖醛酸和乙酸。
(4)糖蛋白细胞壁 盐杆菌属(Halobacterium)细胞壁由糖蛋白组成,包括葡萄糖、葡糖胺、甘露糖、核糖和阿拉伯糖,蛋白质由大量天冬氨酸、谷氨酸等酸性氨基酸组成。这种带强负电荷细胞壁可平衡环境中高浓度 Na+,使其能生活在 20%~25%高盐溶液中。
(5)蛋白质细胞壁少数产甲烷菌的细胞壁是由蛋白质组成的。有的由几种不同蛋白质组成,如甲烷球菌属(Methanococcus)和甲烷微菌属(Methanomicrobium),另一些则由同种蛋白的许多亚基组成,如甲烷螺菌属(Methanospirillum)。
近年来发现,几乎所有古菌的细胞壁都有类结品表面层(S层),由蛋白质或糖蛋白等组成的六角形对称的小单体拼接而成。S层也可直接构成细胞壁。
2.细胞膜:
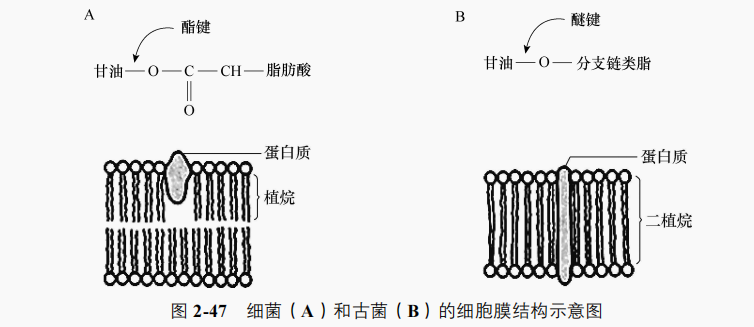
古菌细胞膜与细菌、真核生物有明显差异。①膜中磷脂的亲水性头部仍由甘油组成,但疏水性尾部由长链烃组成,一般是异戊二烯的重复单位(四聚体植烷、六聚体鲨烯等)而不是脂肪酸。②亲水头(甘油)与疏水尾(烃链)通过醚键连接成甘油二醚或二甘油四醚等,细菌及真核生物的疏水尾通过酯键连接甘油和脂肪酸。③古菌细胞膜中有独特的单分子层膜或单、双分子层混合膜。磷脂为二甘油四醚时连接两端两个甘油分子的两个植烷侧链间会发生共价结合形成二植烷,出现独特的单分子层膜(图2-47)。真细菌和真核生物的细胞膜都是双分子层。这类单分子层膜多存在于嗜高温的古菌中,可能是这种膜的机械强度较双分子层膜高。④在甘油分子的 C3 位上可连接多种与真细菌和真核生物细胞膜上不同的基团,如磷酸酯基、硫酸酯基及多种糖基等。⑤古菌细胞膜上含多种独特的脂类,仅在嗜盐菌中就发现细菌红素、α胡萝卜素、β胡萝卜素、番茄红素、视黄醛(可与蛋白质结合成视紫红质)和萘醌等。
3.细胞质:
古菌的细胞质和内含物与细菌的基本相同,如无细胞器、核糖体 70S 等
三、古菌的遗传学特征
古菌的一些遗传学特征与细菌相似,染色体都是单个共价闭合环状 DNA 分子,某些古菌的基因组比一般真细菌的基因组小得多。例如,大肠杆菌的DNA约为 4.6Mb,嗜酸热原体(Thermoplasma acidophilum)的DNA为1.7Mb。詹氏甲烷球菌(Methanococcus jannaschii)的整个基因组已被测序,共有 1738 个基因,约 56%的基因与细菌和真核生物不同。古菌 DNA(G+C)mol%的范围较大,为21%~68%。古菌中只有少数种有质粒。
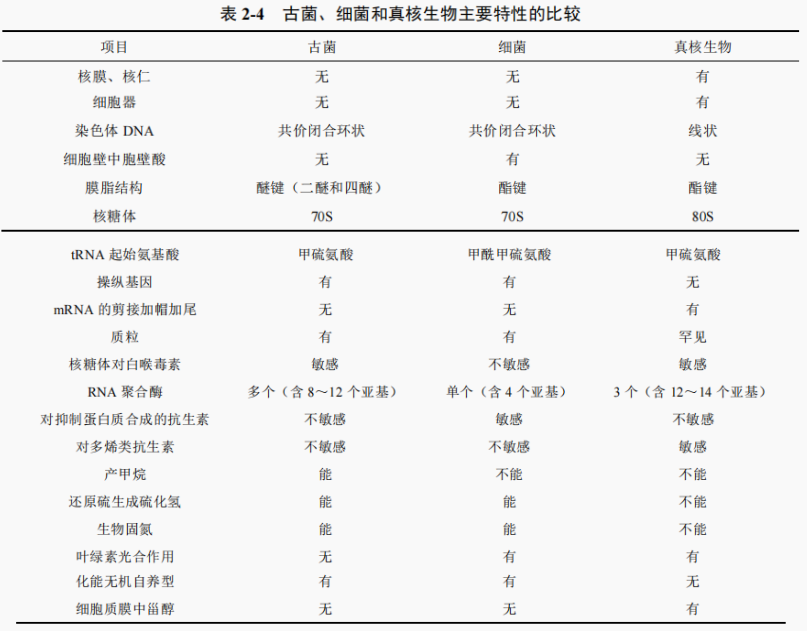
古菌的 RNA 聚合酶类似于真核生物而不同于细菌。古菌的 RNA 聚合酶亚基数8~12 个,比细菌多 4个。古菌的核糖体大小与细菌相同,为 70S,但蛋白质合成开始时的氨基酸与真核生物一样为甲硫氨酸,而细菌是甲酰甲硫氨酸,对抑制蛋白质合成的抗生素的敏感性等均与细菊不同而类似于真核生物。由此可以认为古菌是一类16SrRNA 及其他部分细胞成分在分子水平上与细菌和真核生物有所不同的生物类群。古菌、细菌和真核生物主要特性的比较见表 2-4。
四、古菌的主要类群
1.产甲烷菌这是一类严格厌氧的极端微生物,形态和生理方面差别很大。其共同点是能以氢气、甲酸或乙酸等还原 CO2并产生甲烷。此过程只能在厌氧条件下进行,故它们都是严格厌氧菌,氧气对它们有致死作用。其细胞内不含过氧化氢酶和过氧化物酶,常含有辅酶M(β-巯基乙基磺酸)和能在低电位条件下传递电子的辅酶F420。在荧光显微镜下观察有自发荧光,这是识别产甲烷菌的重要方法。有的能同化 CO2营自养生活,但同化CO2不经卡尔文循环,而是将它直接固定为乙酸盐加以利用。乙酸可以刺激其生长。某些种需要氨基酸、酵母膏、酪素水解物作生长因子。有的可以固氮。产甲烷菌都需要镍、铁、钴等微量元素。
产甲烷菌主要分布在有机质厌氧分解的环境中,如沼泽、湖泊、污水和垃圾处理场、动物的瘤胃及消化道和沼气发酵池中。包括 G+菌和 G-菌,自养和异养,形态有球状、杆状、丝状、螺旋状等多种类型,宽 0.5~1.0μm。细胞壁由假肽聚糖组成。有菌毛,不运动。产甲烷菌根据形态特征、可利用的底物及系统发育分析,目前划分为1门(广古菌门)、3纲(甲烷杆菌纲、甲烷球菌纲、甲烷微菌纲)、5 目(甲烷杆菌目、甲烷球菌目、甲烷微菌目、甲烷八叠球菌目甲烷火菌目)、9科、28属、99种(2000年)。产甲烷菌代谢机制独特,不能利用碳水化合物、蛋白质等复杂有机物,但可利用其他微生物降解有机废物、污水等有机物所产生的乙酸、甲酸、H2和 CO2,等转化为甲烷。沼气发酵既可生产清洁能源和优质有机肥,又可消除污染,还能杀死病菌、虫卵和草籽。
2.嗜盐微生物 嗜盐微生物能在含盐 20%~30%甚至饱和盐水中生活,主要分布在盐湖和晒盐池等高盐度环境中。严格好氧,革兰氏染色阴性,二分裂繁殖,不产生孢子,无休眠状态,细胞呈球状或杆状,(0.5~1.2)μmx(1.0~6.0)μm。大多数不运动,化能有机营养,常以蛋白质、氨基酸等为碳源和能源,一般因具有类胡萝卜素而呈红、橙等颜色。紫膜是嗜盐微生物细胞结构的一个重要特征,除具有光合作用外,还具有光能转换等特性,如将太阳能转换成电能。在厌氧、有光时能合成细菌视紫红蛋白嵌入细胞膜中,利用光能将H+泵出细胞膜,利用由此产生的电化势,在 ATP 酶的催化下,合成 ATP。
嗜盐微生物可用于生产医药和食品中广泛应用的 β-胡萝卜素、胞外多糖、聚 β-羟基丁酸(PHB)、食用蛋白、调味剂、保健食品强化剂、酶保护剂及计算机存储器等,还可用于海水淡化、盐碱地改良及能源开发如太阳能电池等。一些嗜盐微生物可引起食品腐败变质,如副溶血弧菌(Vibrio parahaemolyticus)能污染食品,并引起食物中毒。
根据 16SrRNA序列分析并结合其他生物学性状,将极端嗜盐菌分为盐杆菌属(Halobacte-rium)、盐球菌属(Halococcus)、盐红菌属(Halorubrum)、盐棒菌属(Halobaculum)、富盐菌属(Haloferax)、盐盒菌属(Haloarcula)、嗜盐碱杆菌属(Natronobacterium)和嗜盐碱球菌属(Natronococcus)等26个属。
3.嗜热微生物嗜热微生物简称嗜热菌,是一类依赖于硫、能耐高温(100℃以上)的特殊类群,主要生活在含硫的温泉、泥沼地、火山口、燃烧后的煤矿及含硫的水中。形态有球状、杆状、圆盘状等。绝大多数专性厌氧,以硫作电子受体,进行化能有机营养或化能无机营养的厌氧呼吸产能代谢。嗜热菌可细分为以下5类。

由于生物氧化作用,富硫的热水及其周围环境往往呈酸性,pH 5.0左右,有的低于 1.0。含硫的热泉称硫磺热泉。主要的极端嗜热菌多生活在弱酸性的高热地区,如热网菌(Pyrodictiumsp.)是海洋火山极端嗜热菌中最令人感兴趣的古菌之一,它自 82℃开始生长,最适温度 105℃最高达 110℃,中性条件下利用氢气进行化能无机营养,以硫作电子受体; 火叶菌属(Pyrolobus)在 113℃能生长,121℃可存活1h。已发现有的种能在 121℃下生长,130℃可存活 2h,该菌球形,严格厌氧。极端嗜热菌的呼吸类型多,无论是以有机物、无机物作呼吸底物,还是进行化能有机营养或化能无机营养的厌氧呼吸产能代谢,硫元素在各类呼吸作用中都起关键的作用,可作电子受体和电子供体。加入有机物对其生长有刺激作用。在厌氧条件下进行化能有机营养。许多化能无机营养的极端嗜热菌能利用氢作能源,在好氧条件下以氢作电子供体。
嗜热菌在生产实践和科学研究中有广阔的应用前景,因为嗜热菌具有生长速度快、代谢活动强、产物与细胞的质量比高和培养时不怕杂菌污染等优点,特别是其耐热酶因作用温度高和热稳定性好等突出优点,已在 PCR 等科研和应用领域中发挥越来越重要的作用。有的嗜热菌可用于细菌浸矿、石油和煤炭的脱硫;有助于堆肥、垃圾场等高温环境中有机物的降解;发酵工业中嗜热菌已用于纤维素酶、蛋白酶、淀粉酶、脂肪酶及菊糖酶等多种酶制剂的生产,这些酶制剂热稳定性好,催化反应速度快,易于在室温下保存。
4.嗜冷微生物 它们在 0℃以下也能生长,最适生长温度低于 15℃,最高生长温度低于 20℃。主要生长在雪山、冻土、冰窖、冷藏库等低温环境,在室温中短时间就会死亡。嗜冷微生物在自然界和人工环境的广泛分布反映了其代谢能力的多样性,在自然界的物质循环中起重要作用。许多嗜冷固氮根瘤菌已分离到,一些冷菌能产生降解有机污物等大分子的胞外酶用于污染物处理。嗜冷菌是低温保藏食品发生腐败的主要原因。
5.嗜酸微生物 是能在 pH0.5~4.5 环境中生长的一类微生物,一般分布在酸性矿水、生物滤沥堆、酸性热泉和酸性土壤等酸性环境中,pH5.5 以上不能生长。极端嗜酸微生物是指生长 pH 上限为 3.0、最适生长 pH 在 2.5 以下的微生物。例如,氧化硫杆菌(Thiobacillusthiooxidans)在 pH 低于 0.5的环境中仍能存活,专性自养嗜酸的氧化亚铁硫杆菌(Tferrooxidans)能氧化硫和铁,并产生硫酸,这两种细菌都是极端嗜酸微生物。多年来,一些嗜酸微生物被广泛用于铜等金属的细菌浸矿。也尝试利用硫杆菌分解磷矿粉,提高其溶解度,增加磷矿粉的肥效。用嗜热嗜酸的硫化叶菌(Sulfolobus)脱除煤炭中的硫化物,不仅无机硫化物去除率高,还可去除有机硫化物。一些热嗜酸微生物还是高温酶的来源,已分离出乙醇脱氢酶、β-半乳糖苷酶和苹果酸脱氢酶等耐高温酶,将开拓酶工业的新领域。
6.嗜碱微生物 多数生活在盐碱湖、碱湖、碱池和盐碱土中,环境 pH 可达 11.5 以上最适 pH 8.0~10.0。专性嗜碱微生物可在 pH 11.0~12.0的条件下生长,但在中性条件下却不能生长,如巴氏芽孢杆菌(Bacillus pasteurii)、嗜碱芽孢杆菌(B.alcalophilus)等。大多数嗜碱微生物是好氧菌,有些还是嗜盐菌或中度嗜盐菌。除碱性环境外几乎所有嗜碱芽孢杆菌生长、发芽及芽孢形成都需要 Na+;许多嗜碱菌生长需要多种营养,少数嗜碱芽孢杆菌还能在含甘油谷氨酸、柠檬酸等简单的基础培养基中生长。利用啫碱微生物可生产碱性蛋白酶、碱性淀粉酶碱性果胶酶、碱性纤维素酶等酶制剂。
参考资料
蔡信之, 黄君红, 康贻军. 微生物学 (第四版). 北京: 科学出版社, 2023.
敬请关注“灰藻视界”,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2025-03-03
#创作团队
编制人:木木