探索新鲜水果、蔬菜和食用花卉中的乳酸菌从生物多样性价值化到食物应用
来源:武汉市灰藻生物科技有限公司 浏览量:12 发布时间:2026-07-07 11:04:52
引言
新鲜植物基质(如水果、蔬菜和可食用花卉)中栖息着复杂的微生物群落,这些微生物的生化与生态相互作用对提升食品品质和安全性至关重要。乳酸菌(LAB)在发酵、生物保护和生物活性等方面发挥着关键作用。尽管乳酸菌在食品生物技术中的重要性已广为人知,但其在植物基质中的生态动态与代谢多样性仍缺乏系统研究。理解乳酸菌的多样性及其对基质特异性胁迫的适应机制,对于发掘具有独特技术和生物功能特性的新菌株具有重要意义。
本综述系统总结了近年来关于新鲜植物基质相关乳酸菌的研究成果,重点关注以下方面:
• 鉴定方法:比较了培养依赖性与非培养依赖性方法(如16S rRNA宏条形码、鸟枪法宏基因组学、多组学技术)的应用与局限。
• 功能特性:分析了乳酸菌对恶劣环境的耐受性、微生物间相互作用机制及其主要技术与功能属性。
• 生物技术应用:涵盖植物基发酵食品与饮料开发、益生菌菌种/生物防控方案设计,以及植物副产品的增值利用。
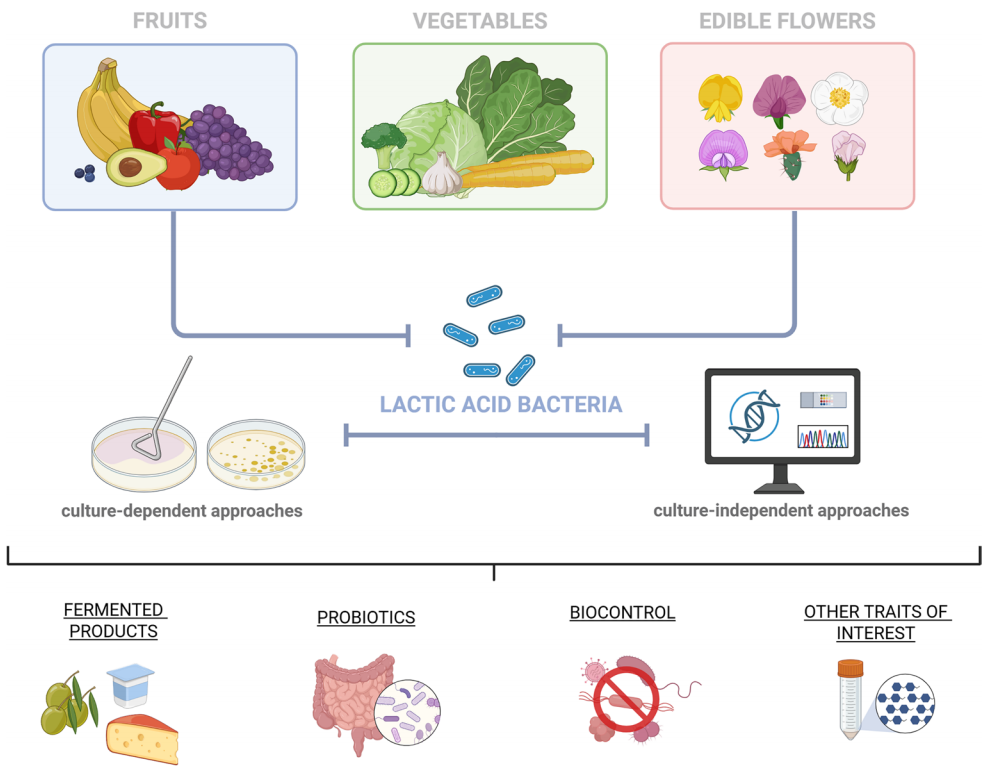
图1. 通过培养依赖性和非培养依赖性策略从水果、蔬菜和可食用花卉中回收乳酸菌的工作流程,及其在食品生物技术中的潜在应用
一、生态背景与微生物群落结构
1.1 植物作为“元生物体”的生态学
植物并非孤立的生物个体,而是与其表面和内部的微生物共同构成了一个“元生物体(Meta-organism)”。在新鲜果蔬和可食用花卉的生态系统中:
• 核心微生物组:无论环境如何波动,总有一部分微生物稳定存在。这部分“常驻居民”对植物的健康和抗病性至关重要。
• 动态平衡:微生物群落的结构受内在因素(植物基因型、组织生理阶段)和外在因素(农业实践、气候、土壤)的双重调控。
• 种植方式的影响:研究表明,有机种植的果蔬(如苹果、蓝莓)往往比常规种植的具有更高的细菌多样性。例如,在常规农业条件下,紫色花卉(Torenia fournieri)中肠杆菌科物种较多,而使用生物堆肥种植的花卉中Granulibacter丰度较低。
1.2 乳酸菌(LAB)在植物界的特殊地位
虽然酵母菌和真菌通常在植物表面占主导地位,但乳酸菌(LAB)是其中不可或缺的“功能型”成员。
• 生存悖论:植物环境对乳酸菌来说是严苛的。与动物肠道或乳制品相比,植物表面具有紫外线辐射强、氧化应激高、蛋白质含量低而糖分(尤其是果糖)丰富的特点。
• 进化适应:为了在植物界生存,乳酸菌进化出了特殊的生理策略。例如,它们能代谢植物特有的糖类,解毒酚类化合物,并表现出对恶劣环境的强耐受性。
• 分类学革新:随着基因组学的发展,乳酸菌属(广义)在2020年经历了重大的重新分类。现在的研究更倾向于使用新属名,如植物乳杆菌属 (Lactiplantibacillus)、乳酸嗜酸菌属 (Lacticaseibacillus) 等,这反映了它们向特定生态位进化的趋势。
• 特别关注:嗜果乳酸菌 (FLAB):这是一个专门适应高果糖环境(如花蜜、成熟水果、蜂蜜)的独特分支,包括蜜蜂乳杆菌属 (Apilactobacillus) 和果糖乳杆菌属 (Fructobacillus)。它们在传粉昆虫的肠道稳态中也扮演着重要角色。
二、研究方法——从培养皿到基因组
为了克服传统方法的局限,现代研究采用了“多相分类法”,结合了培养与非培养技术。
2.1 非培养依赖性方法
这类方法不依赖细菌培养,能更全面地揭示微生物多样性。
○ 16S rRNA 宏条形码技术:
• 原理:通过高通量测序(NGS)特定的16S rRNA基因片段(如V3-V4区),对细菌群落进行属水平或种水平的分析。
• 应用案例:在椰子树液(Kalparasa)的自发发酵研究中,发现新鲜液汁中以明串珠菌属(Leuconostoc)为主(61.15%),而发酵结束时该比例下降,果糖乳杆菌属(Fructobacillus)比例上升。
• 局限:难以区分亲缘关系极近的物种,且PCR扩增可能引入偏差。
○ 鸟枪法宏基因组学 (Shotgun Metagenomics):
• 优势:这是目前最强大的无培养策略。它对环境中所有DNA进行随机测序,不仅能提供菌株水平的分辨率,还能直接预测功能基因(如碳水化合物代谢、抗菌物质合成)。
• 挑战:成本高昂,且难以区分“活细胞”与“死细胞”的DNA。对于乳酸菌这种在植物中丰度较低的菌群,检测难度较大。
○ 多组学整合:
• 结合宏转录组(看基因表达)、宏蛋白质组(看蛋白质合成)和代谢组(看代谢产物),可以全方位解析微生物群落的实际活性。
2.2 培养依赖性方法
乳酸菌是公认的营养需求较高的微生物,通常无法在仅含碳源的最低培养基上生长。通过添加微量营养素、有机化合物、表面活性剂(如吐温80)或调整糖类含量,可有效促进某些乳酸菌的选择性分离。
在厌氧或微需氧条件下,MRS培养基(De Man-Rogosa-Sharpe)是常用的标准培养基,用于回收广谱且代谢多样的乳酸菌,如:
• 明串珠菌属肠膜亚种(Leuconostoc mesenteroides)
• 植物乳杆菌(Lactiplantibacillus plantarum)
• 魏斯氏菌属困惑魏斯氏菌(Weissella confusa)
近年来,越来越多的研究采用表型与分子相结合的策略,从新鲜水果、蔬菜和可食用花卉中分离并鉴定乳酸菌(表1、表2、表3)。
表1. 从生鲜水果中分离的乳酸菌(LAB)菌株汇总,包括基质类型、培养条件、鉴定方法及地理来源
| 分离出的土著乳酸菌物种 | 分离来源 | 培养基和生长条件 | 表征(形态学、生化和分子测试) | 样本采集地点 |
|---|---|---|---|---|
| 推定的 Lb. bulgaricus (保加利亚乳杆菌) | 草莓 | 营养琼脂 - MRS 琼脂 | 通过目视检查和革兰氏染色进行形态学表征 | 印度尼西亚,武吉丁宜,阿耶安格克 (Aie Angek, Tanah Datar district, Indonesia) |
| Lmb. fermentum (发酵乳杆菌), Lpb. plantarum (植物乳杆菌), S. lutetiensis (路氏沙雷菌) | 成熟番石榴,酸浆果 (cape gooseberry) | 在 MRS 肉汤中富集并在 37°C 下培养 1-2 天;涂布于 MRS 琼脂(含 1% CaCO₃)并在厌氧条件下于 37°C 培养 48 小时 | 革兰氏染色和过氧化氢酶试验;16S rRNA 基因扩增(通用引物:FC27 和 RC1492) | 菲律宾,内湖省,洛斯巴ños (Los Baños, Laguna, Philippines) |
| Fb. tropaeoli, Cb. paraalimentarius/kimchii, Lpb. paraplantarum, Lc. lactis, Leuc. pseudomesenteroides, Leuc. citreum, Leuc. mesenteroides, W. cibaria, W. confusa, W. paramesenteroides, W. soli | 木瓜和番茄 | MRS 琼脂 + 放线菌酮 — 37°C 培养 72 小时 | 过氧化氢酶试验,通过 16S 测序进行鉴定;必要时对 pheS 和 recA 基因进行测序。PCR 产物使用特异性引物通过桑格测序。通过 (GTG)₅ rep-PCR 进行种内分型 | 留尼汪岛,法国海外省 (Reunion Island, French overseas department) |
| Lvb. brevis, W. cibaria, Leuc. mesenteroides subsp. dextranicum/mesenteroides, Lc. lactis subsp. lactis, E. faecalis, E. gallinarum/casseliflavus, Fb. tropaeoli, Fb. durionis, Lc. lactis, Lcb. rhamnosus, Pw. fabalis, Leuc. pseudomesenteroides, Leuc. citreum, E. hirae | 番石榴,木瓜,百香果,蛋黄果,欧楂,桑葚,无花果和柿子 | MRS 平板(LAB),fMRS + 2% 果糖(嗜果糖 LAB)。富集:5 mL FYP 肉汤,30°C,24 小时。转接:100 μL FYP 肉汤,培养。FYP 琼脂 + 0.5% CaCO₃。30°C 培养 24–72 小时(LAB,FLAB) | 通过 rep-PCR 基因组指纹图谱对分离株进行分子去重复,并对 16S rRNA 基因的 V1 可变区进行测序,以代表来自不同 LAB 和 FLAB 簇的分离株 | 阿根廷北部,图库曼 (Tucumán, northern Argentina) |
| Fb. pseudoficulneus, Fb. durionis | 苹果、香蕉、冬桃、李子、甜瓜、猕猴桃和荔枝 | FYP肉汤:1% D-果糖,0.5% 蛋白胨,30 °C,24小时,振荡。 接种:50 µL 样品接入含递增果糖浓度(最高30%)的FYP肉汤中。 涂布:含30%果糖+0.5% CaCO₃的FYP琼脂。 进一步培养:在GYP肉汤(葡萄糖)中,30 °C,24小时 | 16 S rRNA基因扩增和测序 | 中国无锡(江苏) |
| W. cibaria | 纳兰吉拉果 (Naranjilla) | 培养:10 g 样品 + 无菌水,室温5天; 接种与培养:MRS琼脂,37 °C 24小时,厌氧条件 | 革兰氏染色,过氧化氢酶,动力,吲哚试验; 从头NGS测序 (Illumina HiSeq X Ten, Macrogen Inc.),PCR扩增 | 苏昆比奥斯,厄瓜多尔 |
| Lc. lactis, Lc. garvieae, W. confusa, W. oryzae, E. faecalis | 野生植物果实 | 直接取样于MRS琼脂,10天; 7 mL 无菌食用油,低氧,厌氧环境; 0.85% 无菌生理盐水稀释,MRS + 1% CaCO₃平板, 37 °C 24小时,厌氧条件 | 16 S rRNA基因扩增(通用引物:27 F 和 1492R)和测序 | 坦布劳,西巴布亚,印度尼西亚 |
| W. paramesenteroides | 人心果,樱桃,香蕉,橙子和李子 | 富集培养:在含0.5% 牛磺胆酸钠和0.5% 低聚半乳糖(或0.5% 低聚果糖或低聚木糖)的MRS-BB肉汤中; 37 °C 48小时, 涂布:MRS平板,37 °C 48小时 | 革兰氏染色和过氧化氢酶试验; 16 S rRNA基因扩增(通用引物:UNI 8 F 和 UNI 1492R)和测序 | 当地市场,印度 |
| E. mundtii (曼氏肠球菌), Leuc. mesenteroides (肠膜明串珠菌) | 青苹果,红苹果,桃子,番石榴,青梨,青番茄,石榴,橙子,柑橘和葡萄 | 在MRS肉汤中富集1 mL,26 °C培养18–24小时,厌氧条件;在MRS平板上重新划线,26 °C培养18小时 | 革兰氏染色,过氧化氢酶测试和碳水化合物发酵谱评估。 16S rRNA基因扩增及测序 | 墨西哥,奇瓦瓦市 |
| 推定且未鉴定的LAB | 葡萄和香蕉 | FYP(1%果糖)和fMRS(MRS+1%果糖);30 °C培养48小时,需氧条件;将选定的菌落重新接种到各自的培养肉汤中,30 °C培养48小时,需氧条件 | 16S rRNA基因扩增及测序 | 爱尔兰,科克郡,费莫伊 |
| Lpb. plantarum (植物乳植杆菌) | 黄皮火龙果 | MRS琼脂 + 苯胺蓝;菌落培养于33和37 °C,48小时,厌氧条件 | 16S rRNA基因扩增及测序 | 未报告 |
| Lvb. brevis (短乳杆菌), Enterococcus sp. (肠球菌属)(通过MALDI-TOF初步鉴定);被鉴定为Lvb. brevis的最佳益生菌候选菌株 | 苹果,香蕉,葡萄和橙子 | MRS琼脂,37 °C,48小时,厌氧条件 | 革兰氏染色,过氧化氢酶测试和运动性评估,使用MALDI-TOF初步鉴定,16S rRNA基因扩增(通用引物:341 F和806R)及测序 | 巴西,帕拉伊巴州,若昂佩索阿 |
| Lpb. plantarum (植物乳植杆菌) 和 Lmb. fermentum (发酵乳杆菌) | 樱桃番茄,蓝莓,黑莓,樱桃和苹果 | 在MRS肉汤中富集,37 °C,24小时,厌氧,涂布于MRS琼脂,37 °C,48小时,厌氧条件 | 革兰氏染色,过氧化氢酶测试,碳水化合物发酵测试,16S rRNA基因扩增及测序 | 中国,丽水,南京及南京当地市场 |
| Lpb. plantarum (植物乳植杆菌) | 香蕉,木瓜,菠萝和橙子 | 涂布于MRS琼脂,30 °C,48小时,厌氧条件 | 革兰氏染色,过氧化氢酶测试,CO₂产生测试,16S rRNA基因扩增及测序 | 喀麦隆西部,门ou瓦省 (Menoua Division),杜尚 (Dschang) |
| P. pentosaceus (戊糖片球菌) | Mandacuru (一种仙人掌果) | 在MRS肉汤中富集,37 °C,72小时,100 rpm;涂布于MRS琼脂,37 °C,48小时,厌氧条件 | 革兰氏染色,过氧化氢酶试验;16S rRNA基因扩增及测序 | 喀麦隆 |
| Lpb. plantarum (植物乳杆菌) 和 P. pentosaceus (戊糖片球菌) | 巴西莓 (Açaí fruits) | 涂布于MRS琼脂,37 °C,48小时,厌氧条件 | 革兰氏染色,过氧化氢酶试验,细胞色素氧化酶活性评估;16S rRNA基因扩增及测序 | 巴西帕拉州(PA)和马兰昂州(MA)的Combu岛、贝伦、阿拜图巴、布雷维斯、圣塔伦及Zé Doca |
| Lpb. plantarum (植物乳杆菌) | 香蕉 | 在MRS肉汤中富集,30 °C,24小时,厌氧;在含0.5% CaCO3的MRS上三个浓度梯度培养,48小时,厌氧条件 | 革兰氏染色,过氧化氢酶试验,葡萄糖产气,明胶液化试验,乳酸异构体测定,糖发酵(API 50 CH);16S rRNA基因扩增(通用引物:27 F 和 1492R)及测序 | 中国海南五指山香蕉公园 |
| E. faecium (屎肠球菌), E. durans (耐久肠球菌), E. lactis (乳肠球菌), P. acidilactici (乳酸片球菌) | 番茄、草莓和桃子 | MRS琼脂涂布,厌氧条件,37 °C,48小时,CO2培养箱,50%甘油和50%水保存菌种 | 革兰氏染色和过氧化氢酶试验;16S rRNA基因扩增(通用引物:27 F 和 1492R)及测序 | 阿拉伯联合酋长国沙迦 |
| Limosilactobacillus sp. (发酵乳杆菌属) | 诺丽果 | 在MRS肉汤中富集,37°C,24小时,厌氧条件;划线接种于MRS琼脂,37°C,厌氧条件 | 通过目视检查进行形态学表征,革兰氏染色,过氧化氢酶试验,16S rRNA基因扩增(通用引物:27 F和1392R)及测序 | 印度希瓦莫加 (Shivamogga) |
| E. faecium (屎肠球菌) | 蓝樱桃 | 在MRS肉汤中富集24小时(30°C),并在37°C下重新培养24小时,厌氧条件 | 16S rRNA基因扩增(通用引物:27 F和1392R)及测序 | 印度迈索尔 (Mysuru) |
| Leuc. mesenteroides (肠膜明串珠菌) | 香瓜茄 (Pepino) | MRS + 蔗糖 (5%),48小时或30°C,需氧条件 (110 rpm) | 未明确报道 | 中国山西临汾 |
| Lpb. plantarum (植物乳杆菌) | 榕树和 Amrutha balli (Tinospora cordifolia) | 在MRS肉汤中富集,37°C培养48小时,厌氧条件 | 16S rRNA基因扩增(通用引物:8 F和1391)及测序 | 印度卡纳塔克邦 (迈索尔和曼迪亚区) |
| Lvb. brevis (短乳杆菌) | 香蕉 | MRS琼脂,37°C培养24–48小时,厌氧条件 | 革兰氏染色,过氧化氢酶试验,产酸产气评估,16S rRNA基因扩增(通用引物:16SrRNA-F和16SrRNA-R) | 印度赖丘尔 (Raichur) |
| Lpb. plantarum (植物乳杆菌) | 野生植物样本:木糖醇、芦荟、角豆、桑葚和草莓树 | MRS琼脂 + 1.5% CaCO₃;37°C培养48小时,需氧条件 | 使用通用引物寡核苷酸 (BSF8 和 BSR1541) 进行16S rRNA基因扩增 | 意大利普利亚 (Apulia) |
| Lpb. argentoratensis (阿根廷乳杆菌) | 菠萝蜜 | 在MRS肉汤中富集,37°C培养24小时;划线接种于MRS,37°C培养24小时 | 革兰氏染色,形态学评估和过氧化氢酶试验。16S rRNA基因扩增(通用引物:27 F和1392R)及测序 | 印度希瓦莫加 (Shivamogga) |
| Lcb. rhamnosus (鼠李糖乳杆菌), Lcb. paracasei (副干酪乳杆菌), Lcb. casei (干酪乳杆菌), Lnb. buchneri (布氏乳杆菌), Lpb. plantarum (植物乳杆菌), Lvb. brevis (短乳杆菌) | 蓝莓 | MRS琼脂 + CaCO₃ (2%),37°C培养24–72小时,厌氧条件 | 通过目视检查进行形态学表征,革兰氏染色和过氧化氢酶试验,16S rRNA基因扩增(通用引物:27 F和1492R) | 中国黑龙江省黑河市 |
| E. faecalis (粪肠球菌) | 番茄和白桑葚 | 划线接种于FYP,37°C培养24小时,需氧条件 | 革兰氏染色和过氧化氢酶试验。16S rRNA基因扩增(通用引物:27 F和1492R) | 土耳其安卡拉卡赫拉曼卡赞 (Kahramankazan) |
| 尚未鉴定 | 刺梨,ñangapirí,chañar,tutá | MRS琼脂 | 革兰氏染色,过氧化氢酶试验和芽孢形成评估; | 南美洲大查科 (Gran Chaco),阿根廷 |
表2. 从蔬菜中分离的乳酸菌(LAB)菌株汇总,包括基质类型、培养条件、鉴定方法及地理来源
| 分离出的本土乳酸菌(LAB)种类 | 分离来源 | 培养基和生长条件 | 表征(形态学、生化和分子测试) | 样本采集地点 |
|---|---|---|---|---|
| Lpb. paraplantarum(旁植物乳杆菌) | 木瓜、山药、芋头、甘蔗和木薯叶 | 在 MRS 肉汤中富集,在 MRS 琼脂上于 37°C 厌氧培养 48 小时 | 革兰氏染色和过氧化氢酶测试,16S rRNA 基因扩增(通用引物:27 F 和 1492R) | 台湾屏东 |
| E. faecalis(粪肠球菌), Lpb. plantarum(植物乳杆菌), W. paramesenteroides(副肠膜明串珠菌) P. pentosaceus(戊糖片球菌), E. hirae(赫氏肠球菌)和其他推定的未鉴定 LAB | 韭葱、甜叶菊和欧芹 | 在 MRS 肉汤中富集并于 37°C 培养 1-2 天;涂布于 MRS 琼脂(含 1% CaCO₃)并于 37°C 厌氧培养 48 小时 | 革兰氏染色和过氧化氢酶测试;16S rRNA 基因扩增(通用引物:FC27 和 RC1492) | 菲律宾 Laguna 省 Los Baños |
| Lpb. plantarum(植物乳杆菌), Lactobacillus sp.(乳杆菌属), Weissella sp.(魏斯氏菌属), Enterococcus sp.(肠球菌属) | 花椰菜、小黄瓜、四季豆、葫芦巴、豇豆、苦瓜、四季豆、番茄、棱角丝瓜和瓠瓜,以及黄瓜 | 在 MRS 肉汤中于 30°C 富集并培养 48 小时(微需氧条件) | 使用 Biolog 进行代谢分析,使用 MALDI-TOF MS 进行蛋白质组分析以及 16S rRNA 基因测序 | 印度 Junnar, Ambegaon 和 Khed |
| P. pentosaceus(戊糖片球菌), W. confusa(混淆魏斯氏菌), Lpb. plantarum(植物乳杆菌) | 黄瓜、生菜和卷心菜 | 涂布于 MRS 琼脂(条件未清楚报告) | 革兰氏染色、过氧化氢酶和氧化酶测试,孢子形成评估,16S rRNA 基因扩增 | 尼日利亚 |
| E. mundtii(蒙德肠球菌), E. faecium(屎肠球菌), Leuc. mesenteroides(肠膜明串珠菌) | 奇拉卡辣椒、墨西哥辣椒、玉米、西葫芦、生菜、黄瓜、辣椒和豆芽 | 在 MRS 肉汤中于 26°C 厌氧富集 18-24 小时,并在 26°C 的 MRS 琼脂平板上培养 18 小时 | 革兰氏染色、过氧化氢酶测试、碳水化合物发酵谱评估,16S rRNA 基因扩增(通用引物:27 F 和 1492R) | 墨西哥奇瓦瓦市 |
| 未鉴定的 LAB | 花椰菜和菠菜 | FYP(1% 果糖)和 fMRS(MRS + 1% 果糖),在 30°C 需氧条件下培养 48 小时 | 16S rRNA 基因扩增 | 爱尔兰科克郡 Fermoy |
| Lpb. plantarum (植物乳杆菌) | 芝麻菜和生菜 | 37°C MRS 琼脂培养 48 小时 | 16S rRNA 基因扩增 (通用引物: 27 F 和 1492R) | 葡萄牙波尔图 |
| Leuc. mesenteroides (肠膜明串珠菌), W. soli (土壤魏斯氏菌) | 胡萝卜 | 30°C MRS 平板划线,厌氧条件下培养 24–48 小时 | 16S rRNA 基因扩增 (通用引物: 8 F 和 1492R) | 意大利阿布鲁佐奥尔图基奥 |
| E. faecium (屎肠球菌), E. durans (耐久肠球菌), E. lactis (乳肠球菌), P. acidilactici (乳酸片球菌) | 黄瓜、生菜、欧芹和卷心菜 | 37°C MRS 琼脂,厌氧条件下培养 24 小时 | 革兰氏染色和过氧化氢酶试验;16S rRNA 基因扩增 (通用引物: 27 F 和 1492R) | 阿联酋沙迦 |
| Lpb. plantarum (植物乳杆菌) | 西兰花小花 | 37°C MRS 肉汤富集培养 24 小时 | 通过目视检查进行形态学表征,革兰氏染色,16S rRNA 基因扩增 (通用引物: 8 F 和 16SR) | 中国东莞 |
| Lpb. plantarum (植物乳杆菌), Lb. helveticus (瑞士乳杆菌), Lgb. salivarius (唾液乳杆菌), Lb. acidophilus (嗜酸乳杆菌) | 甜菜根汁 | 37°C MRS 肉汤富集培养 24 小时,并在 37°C MRS 琼脂平板上培养 48 小时 | 革兰氏染色,过氧化氢酶活性,葡萄糖产 CO2,糖发酵评估(蔗糖、甘露醇、鼠李糖、山梨醇和麦芽糖),16S rRNA 基因扩增(通用引物) | 伊朗马什哈德 |
| 推定 Lpb. argentoratensis (斯特拉斯堡乳杆菌) | 胡萝卜、萝卜和黄瓜 | 未报道 | 通过目视检查进行形态学表征,革兰氏染色,16S rRNA 基因扩增 | 印度普拉亚格拉吉(阿拉哈巴德) |
| Lvb. brevis (短乳杆菌), Lc. lactis (乳酸乳球菌) | 龙舌兰汁液 | 未报道分离方案。 在37 °C的MRS肉汤中复壮菌株 | 未报道 | 墨西哥 |
| Leuc. mesenteroides ssp. mesenteroides (肠膜明串珠菌), Leuc. mesenteroides ssp. dextranicum (葡聚糖明串珠菌), Lc. garvieae (加氏乳球菌), Leuc. citreum (柠檬色明串珠菌), Leuc. carnosum (肉明串珠菌) | 花椰菜和西兰花 | 在30°C的MRS肉汤中富集48小时,并在MRS琼脂平板上培养 | 革兰氏染色,使用VITEK®2系统确认鉴定 | 伊拉克巴格达市场 |
| Ltb. curvatus (弯曲乳杆菌), Lpb. plantarum (植物乳杆菌), E. casseliflavus (卡氏肠球菌), P. acidilactici (乳酸片球菌), Leuc. mesenteroides (肠膜明串珠菌), Lvb. brevis (短乳杆菌), W. cibaria (魏斯氏菌), P. pentosaceus (戊糖片球菌), Ltb. sakei (肉糖乳杆菌) | 红椒和青椒 | 在厌氧/微需氧条件下,于37°C的MRS琼脂上培养48小时 | 显微形态观察,革兰氏染色,过氧化氢酶试验。 通过带有特定引物(GTG)5的rep-PCR进行菌株水平表征 | 阿根廷北部图库曼 |
| Lcb paracasei ssp. paracasei (副干酪乳杆菌), Lc. lactis ssp. lactis (乳酸乳球菌), Lvb. brevis (短乳杆菌), Lcb. rhamnosus (鼠李糖乳杆菌), Lpb. pentosus (戊糖乳杆菌) | Üçburun辣椒 | 在MRS上培养:i) 37°C 24小时(需氧和厌氧条件)以及 ii) 30°C 72小时(需氧条件) | 革兰氏染色,过氧化氢酶试验,氧化酶试验,CO2产生试验,碳水化合物发酵谱评估(API 50 CH试剂盒),M17、BHI、NA、MRS上的生长评估,通过50 CH API软件鉴定 | 土耳其 |
| Lnb kosonis (科斯乳杆菌), Lnb. curieae (居里乳杆菌) | 菊芋 | 在37°C的FYP或IVP中富集72小时,并在需氧条件下于37°C培养 | 显微形态观察,革兰氏染色以及过氧化氢酶和氧化酶试验,16 S rRNA扩增 | 以色列耶路撒冷 |
| Lpb. plantarum (植物乳杆菌) | 橄榄皮 | 在MRS琼脂上培养 | 显微形态观察,革兰氏染色,过氧化氢酶和糖发酵(API 50 CHL)试验,16 S rRNA基因扩增(通用引物:27 F 和 1490R) | 意大利卡塔尼亚 |
表3. 从食用花卉中分离的乳酸菌(LAB)菌株汇总,包括基质类型、培养条件、鉴定方法及地理来源
| 分离出的本土乳酸菌(LAB)物种 | 食用花卉 | 培养基和生长条件 | 表征(形态学、生化和分子测试) | 样本采集地点 |
|---|---|---|---|---|
| Fb. fructosus (果糖乳杆菌), Apb. kunkeei (昆氏阿波乳酸菌) | 水仙、粉红玫瑰、红玫瑰、黄玫瑰和向日葵 | FYP + 果糖 (1% – 30%),30°C 培养 24 小时 | 形态学观察,16S rRNA 扩增,碳水化合物发酵谱评估(API 50 CH 试剂盒) | 无锡,(江苏),中国 |
| E. durans (耐久肠球菌), Lc. lactis (乳酸乳球菌), Enterococcus sp. (肠球菌属), E. lactis (乳肠球菌), Lpb. plantarum (植物乳杆菌) | 栀子花,木槿 (Hibiscus syriacus),龙葵 (Solanum torvum),银合欢 (Leucaena leucocephala) | 样品在 MRS 肉汤中富集,并在厌氧条件下 37°C 培养 48–72 小时 | 革兰氏染色,过氧化氢酶测试和定性形态学鉴定;16S rRNA 扩增 | 那空是贪玛叻 (Nakhon Si Thammarat),泰国 |
| Lvb. brevis (短乳杆菌), W. cibaria (食威氏菌), Leuc. mesenteroides subsp. mesenteroides (肠膜明串珠菌), Leuc. pseudomesenteroides (假肠膜明串珠菌), Lc. lactis subsp. lactis (乳酸乳球菌), E. faecalis (粪肠球菌), E. gallinarum/casseliflavus (鸡肠球菌/卡氏肠球菌), E. casseliflavus (卡氏肠球菌), Fb. tropaeoli (旱金莲乳杆菌), Fb. durionis (榴莲乳杆菌), Lc. lactis (乳酸乳球菌), Leuc. citreum (柠檬明串珠菌), E. hirae (赫氏肠球菌) | 木瓜花,百香果花,枇杷花,苹果花 | MRS 平板 (LAB),fMRS + 2% 果糖 (嗜果糖 LAB)。 富集:5 mL FYP 肉汤,30°C,24 小时。复壮:100 µL FYP 肉汤,接种于 FYP 琼脂 + 0.5% CaCO3。培养 30°C,24–72 小时 (LAB, FLAB) | 通过 rep-PCR 基因组指纹图谱对分离株进行分子去重复,并对来自不同 LAB 和 FLAB 簇的代表性分离株的 16S rRNA 基因的 V1 可变区进行测序 | 图库曼 (Tucumán),阿根廷北部 |
| Leuc. mesenteroides (肠膜明串珠菌), Leuc. mesenteroides subsp. jonggajibkimchii (钟家吉泡菜明串珠菌), Fb. fructosus (果糖乳杆菌), Lc. lactis (乳酸乳球菌), E. faecalis (粪肠球菌), Lpb. plantarum (植物乳杆菌) | 切花 | 在 FYP (1% 果糖) 和 fMRS (MRS + 1% 果糖) 中培养;30°C 需氧培养 48 小时;菌落转移至各自的肉汤培养基;30°C 需氧培养 48 小时 | 16S rRNA 基因扩增和测序 | 基尔沃思郡 (Kilworth, County Cork),爱尔兰 |
| 未鉴定的球菌状 LAB | 仙人掌花 (Nopal flower) | 在 MRS 肉汤中富集;26°C 厌氧培养 18–24 小时;在 MRS 琼脂上传代培养;26°C 培养 18 小时 | 革兰氏染色,过氧化氢酶测试,糖类发酵评估 16S rRNA 基因扩增(通用引物:27 F 和 1492R)和测序 | 奇瓦瓦市 (Chihuahua City),墨西哥 |
| Fb. fructosus (果糖乳杆菌), Apb. kunkeei (昆氏蜜蜂球菌), E. durans (耐久肠球菌), E. faecium (屎肠球菌), Lvb. brevis (短乳杆菌) | Hibiscus rosa-sinensis (朱槿), Rosa rugosa (玫瑰), Tagetes erecta (万寿菊) | MRS肉汤富集培养,24小时,30 °C;沉淀回收,重悬于MRS肉汤;涂布于MRS琼脂,30 °C培养72小时 | 16S rRNA基因扩增(通用引物:27 F 和 1492R)及测序 | 伊拉克,巴格达,Al-Jadriya |
| Lpb. plantarum (植物乳杆菌) | 香蕉花 | MRS肉汤富集培养,24小时,30 °C,厌氧;三种浓度梯度在含0.5% CaCO₃的MRS上培养,48小时,厌氧 | 革兰氏染色,过氧化氢酶试验,葡萄糖产气,明胶液化试验,乳酸异构体测定,糖发酵试验 (API 50 CH), 16S rRNA基因扩增(通用引物:27 F 和 1492R)及测序 | 五指山香蕉园 (中国海南) |
| E. faecium (屎肠球菌), Lpb. plantarum (植物乳杆菌), Lcb. paracasei (副干酪乳杆菌) | 薰衣草根际 | 在营养琼脂上培养并在 28° C 下孵育 | 革兰氏染色,通过 MALDI-TOF MS 进行细菌鉴定;获取蛋白谱;与 MALDI Biotyper CA 数据库进行光谱比对以进行属/种水平的鉴定 | 土耳其,安卡拉 |
2.2.1 乳酸菌的分类学更新
近年来,基因组测序和系统发育分析促使传统广义乳杆菌属被重新整合为多个功能特化的新属,包括:
• 植物乳杆菌属(Lactiplantibacillus)
• 乳酸嗜酸菌属(Lacticaseibacillus)
• 莱维乳杆菌属(Levilactobacillus)
• 利莫乳杆菌属(Limosilactobacillus)
• 共伴乳杆菌属(Companilactobacillus)等
这种重新分类揭示了各属在基因组成、代谢途径和适应策略上的显著差异。
代表性物种与分布:
• 植物乳杆菌(Lpb. plantarum)代谢灵活、基因组可塑性强,广泛定殖于野生植物、花朵、热带水果等。
• 乳酸嗜酸菌属(Lacticaseibacillus)如 Lcb. rhamnosus、Lcb. paracasei,见于无花果、蓝莓、辣椒等。
• 短乳杆菌(Lvb. brevis)可定殖于番石榴、番荔枝花、龙舌兰汁液、橙子等多样化生态位。
• 肠球菌属与魏斯氏菌属营养偏好相似,表型特征重叠,常需全基因组测序或MALDI-ToF MS进行准确鉴定。
MALDI-ToF MS(基质辅助激光解吸/电离飞行时间质谱)已被证明在快速、可靠地鉴别菌株方面特别有效,尤其是与16S rRNA基因分析联合使用时。
2.2.2 嗜果乳酸菌(FLAB)
FLAB是乳酸菌中一个独特的系统发育分支,仅包含:
• 蜜蜂乳杆菌属(Apilactobacillus)
• 果糖乳杆菌属(Fructobacillus)
FLAB包括:
• 专性果糖嗜好者,如 Apilactobacillus kunkeei、A. apis、A. micheneri;
• 兼性果糖嗜好者,如部分 Lvb. brevis 菌株。
它们表现出特殊的基因组适应特征:
• 基因组缩小
• GC含量降低
• adhE 基因部分缺失
• 需外源电子受体再生NAD⁺
分布:已从新鲜花卉、切花及木瓜、无花果、柿子等富糖果实中分离出 Fructobacillus tropaeoli、Fb. durionis、Fb. fructosus 和 Apilactobacillus kunkeei。Sakandar等通过逐步提高果糖浓度(1%–30%),在香蕉和桃子中检测到 Fb. pseudoficulneus。
FLAB的演化为生态专化驱动适应性进化提供了典型范例。理解这一动态过程,对筛选高价值菌株和开发创新生物技术应用具有重要意义。
三、生物技术应用——从生态到产业
植物源乳酸菌不仅是发酵剂,更是多功能的生物资源,具有巨大的应用潜力。
3.1 食品发酵与功能化
• 植物基食品革新:随着植物基饮食的流行,乳酸菌被用于发酵燕麦、大豆、杏仁等植物基底。例如,从可食用花卉中分离的菌株能显著酸化燕麦饮料(pH降至3-4),赋予其独特的风味。
• 营养提升:研究表明,利用特定的植物乳杆菌(Lpb. plantarum)发酵蓝莓汁,能显著提高酚类物质含量和抗氧化活性(最高提升107.9%)。
• 质地改良:乳酸菌产生的胞外多糖(EPS)是天然的增稠剂和稳定剂。在植物基饮料中,EPS可以模拟乳制品的口感,解决植物蛋白饮料“水感”过重的问题。
3.2 生物保护与采后保鲜
• 天然防腐剂:乳酸菌产生的有机酸(乳酸、乙酸)、细菌素(如类似肠溶素A的物质)能有效抑制腐败菌。
• 抗真菌应用:灰霉病(Botrytis cinerea)是草莓等水果采后腐烂的主要原因。研究证实,植物乳杆菌(Lpb. plantarum CM-3)处理草莓,可将灰霉病发生率降低高达75%。
3.3 益生菌与后生元特性
• 健康潜力:从可食用花卉和植物中分离的某些菌株(如E. durans, Lpb. plantarum)表现出良好的胃肠道耐受性(耐酸、耐胆盐)。
• 功能性测试:部分菌株具有胆固醇吸收能力、胆盐水解酶(BSH)活性,甚至在体外模型中显示出免疫调节和抗炎作用。
• 安全性评估:对于拟用于食品的菌株,必须进行严格的安全性评估,包括检测溶血活性、抗生素耐药基因及生物胺生成能力。
3.4 副产品增值与青贮饲料
• 循环经济:利用乳酸菌发酵非商品级或受损的果蔬(如香蕉、木瓜),将其转化为高价值的动物饲料(青贮)。
• 案例:使用植物乳杆菌(Lpb. plantarum CG1)发酵非商品香蕉,配合单宁酶和蔗糖,可以生产出营养价值更高的青贮饲料。这不仅减少了农业废弃物,还缓解了饲料短缺问题。
四、总结与未来展望
4.1 当前面临的挑战
• 知识空白:目前国际微生物库中,植物来源的乳酸菌株代表性严重不足。
• 技术限制:植物基质成分复杂(如多酚、纤维素),干扰DNA提取和PCR反应;且乳酸菌在植物表面密度低,增加了检测难度。
4.2 未来的研究方向
✅ 精准设计发酵剂:不再盲目套用乳制品发酵菌种,而是根据植物基质的特定成分(如糖谱、缓冲能力),理性设计适应性更强的发酵剂。
✅ 深度基因组学:利用泛基因组分析(Pan-genome analysis)和基因-性状关联研究,绘制乳酸菌在植物生态系统中的适应性图谱。
✅ 体内模型验证:从体外实验转向体内模型(如秀丽隐杆线虫、小鼠、斑马鱼),更真实地验证其益生功能和安全性。
参考文献
1.Abedin MM, Chourasia R, Phukon LC et al (2024) Lactic acid bacteria in the functional food industry: biotechnological properties and potential applications. Crit Rev Food Sci Nutr 64:10730–10748. https://doi.org/10.1080/10408398.2023.2227896
2.Bal M, Önlü H, Osmanağaoğlu Ö (2024) Isolation and In-Vitro Probiotic Characterization of Fructophilic Lactic Acid Bacteria from Different Plants and The Digestive System of Bees. J Inst Sci and Tech 14:1013–1030. https://doi.org/10.21597/jist.1441350
3.Chen Y, Sun J, Luo M et al (2024) Quantifying the viability of lactic acid bacteria using ratiometric fluorescence assays. Microchem J 206:111485. https://doi.org/10.1016/j.microc.2024.111485
4.de Vasconcelos Medeiros GKV, Martins ACS, Vasconcelos MG et al (2024) Cereus jamacaru DC. (mandacaru) fruit as a source of lactic acid bacteria with in vitro probiotic-related characteristics and its protective effects on Pediococcus pentosaceus during lyophilization and refrigeration storage. Int J Food Microbiol 417:110695. https://doi.org/10.1016/j.ijfoodmicro.2024.110695
5.Du S, Xu L, Jiang C, Xiao Y (2025) Novel strategy to understand the bacteria-enzyme synergy action regulates the ensiling performance of wheat straw silage by multi-omics analysis. Int J Biol Macromol 289:138864. https://doi.org/10.1016/j.ijbiomac.2024.138864
6.Ibrahim HA, Khelkal IN, Muhsin YM (2024b) Improvement of Bacteriocin Production by Natural Zeolite and Detection of Antibacterial Activity of Leuconostoc carnosum Purified Carnosin. Basrah J Agric Sci 37:37–53. https://doi.org/10.37077/25200860.2024.37.2.4
相关产品
HZB357425:植物乳杆菌 | Lactobacillus plantarum
HZB358700:粪肠球菌 | Enterococcus faecalis
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-06-19
编制人:思琪
审稿人:叶凡