噬菌体内溶素在食品安全领域的应用综述:进展、挑战与展望
来源:武汉市灰藻生物科技有限公司 浏览量:17 发布时间:2026-06-30 15:59:32
摘要
食源性疾病是食品安全领域的重要挑战,主要由大肠杆菌、沙门氏菌、金黄色葡萄球菌、单核细胞增生李斯特菌、弯曲杆菌和梭菌等致病菌污染食品所致。
内溶素作为噬菌体编码的肽聚糖水解酶,能通过裂解细菌细胞壁发挥抗菌作用,尤其对革兰氏阳性菌效果显著。
近年来,内溶素因其高选择性、独特作用机制、易于工程化改造及不易诱导耐药性等优势而备受关注。
本文综述了内溶素在清除食源性致病菌生物被膜、快速检测病原菌等方面的应用,总结了其在畜牧业和各类食品基质中的实践效果及作为膳食补充剂的局限性。
此外,本文重点介绍了靶向革兰氏阴性菌的工程化内溶素开发新技术。研究表明,内溶素对食源性致病菌安全有效,且对人类细胞和有益微生物群无不良影响,有望作为一种功能性生物防腐剂,在保障食品安全的同时保持食品原有风味。
一、综述
食源性致病菌导致的疾病和死亡已成为全球性公共卫生问题。据世界卫生组织统计,全球每年约6亿人因食用污染食品而患病,约42万人死亡,造成约1100亿美元的经济损失。致病菌可在食品生产、运输、处理和包装等各环节造成污染,而现有防控手段存在诸多不足:
• 物理方法:高压处理可能破坏食品质地,辐照可能影响感官品质。
• 化学防腐剂:醋酸、次氯酸盐等化学物质会在食品中留下有害残留,消费者接受度低。
• 抗生素:畜牧业中广泛使用抗生素导致微生物群紊乱,并加速了多重耐药菌的产生。
因此,亟需开发新型抗菌技术。噬菌体源内溶素因其独特的抗菌优势——包括对耐药菌有效、裂解迅速、靶向特异、耐药风险低等——被认为是极具前景的天然防腐剂。本文将从内溶素的结构与分类、关键特性、应用实践、工程化改造及面临挑战等方面进行系统综述。
1.1 内溶素的作用机制
细菌裂解是一个三步模型系统,涉及三种蛋白质的协同作用:
• 穴蛋白(Holin):在细菌内膜上形成小孔。
• 内溶素(Endolysin):通过小孔进入周质空间,降解肽聚糖层。
• 跨度蛋白(Spanin):破坏外膜,完成裂解过程。
对于革兰氏阴性菌,还存在另一种途径——小孔穴蛋白-分泌型激活调控内溶素途径。过去认为仅破坏内膜和肽聚糖即可导致裂解,但最新研究表明,跨度蛋白介导的外膜破坏同样不可或缺。
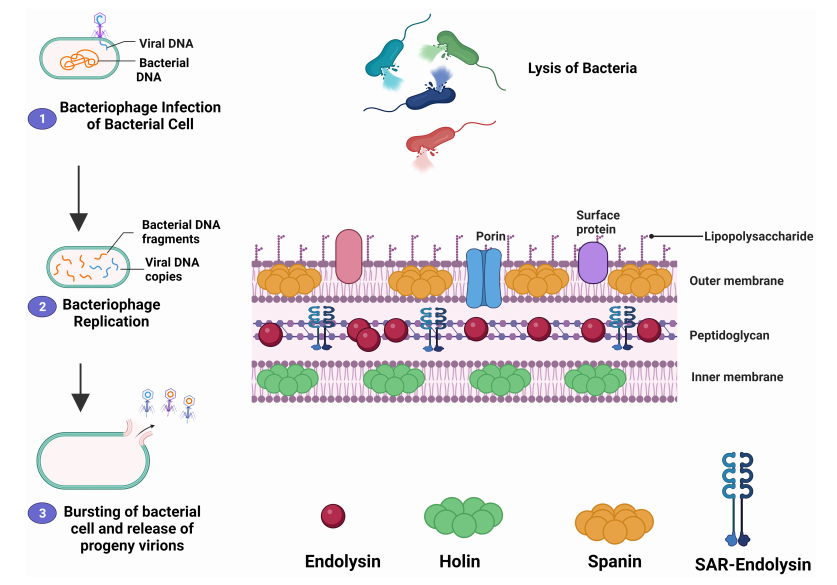
图1、示意图展示了噬菌体裂解周期及噬菌体衍生内溶素的作用机制
注:该图展示了穿孔素-内溶素系统如何到达并穿透肽聚糖层,进而裂解细菌。该机制表明,在第一步中,噬菌体通过其特异性的尾丝蛋白识别并结合到细菌特异性的细胞受体上。附着后,噬菌体通过尾部将其遗传物质注入细菌体内,随后组装形成子代噬菌体。穿孔素(holin)会形成通道,使内溶素(endolysin)得以进入并输送至细菌的肽聚糖层。在最后一步,细菌细胞通过内溶素的作用发生降解。
1.2 内溶素的结构
模块化结构:大多数内溶素由N端酶活性结构域(EAD)和C端细胞壁结合结构域(CBD)通过柔性连接区相连而成。EAD负责切割肽聚糖键,CBD负责识别并结合靶标细胞壁。感染革兰氏阳性菌的噬菌体内溶素多属此类。
球状结构:作用于革兰氏阴性菌的内溶素通常仅有球状EAD,不含CBD;但也有研究发现部分革兰氏阴性内溶素在N端含有一个或两个CBD。
二、内溶素的主要特征
2.1 广谱裂解活性
内溶素对缺乏外膜的革兰氏阳性菌效果最佳,但对革兰氏阴性菌也具有一定活性。其特异性通常处于属或种水平,由CBD决定。部分内溶素具有广谱活性:
• Pal:可清除15种肺炎链球菌血清型,包括青霉素耐药株。
• phi11:对多种葡萄球菌有效,如金黄色葡萄球菌、表皮葡萄球菌等。
• LysAB54、LysP53:对鲍曼不动杆菌、铜绿假单胞菌等多重耐药革兰氏阴性菌有效。
• Abtn-4、LysSS:兼具抗革兰氏阳性和阴性菌的广谱活性。
2.2 不易产生耐药性
内溶素作用于细菌细胞壁表面的肽聚糖,而多数耐药机制发生在胞内,故细菌难以对内溶素产生耐药性。实验表明,肺炎链球菌、芽孢杆菌和MRSA均未对内溶素产生耐药,而相同条件下对抗生素却产生了耐药。目前尚无细菌对内溶素耐药性的公开报道。
2.3 对正常微生物群无害
与广谱抗生素不同,内溶素能选择性杀灭特定致病菌,对有益菌群影响极小:
• PlyV12:对耐万古霉素肠球菌有效,不转移耐药基因。
• S25-3:在抗金黄色葡萄球菌的同时,增加了小鼠皮肤微生物多样性。
• Pal:不影响人咽部常见微生物。
2.4 安全性良好
多项研究证实内溶素具有良好的安全性:
• 免疫原性:Pal和Cpl-1虽可引起IgG升高,但IgE水平低,过敏风险小。
• 动物实验:PlyCD1-174、SAL200等在小鼠、犬类中未见明显不良反应。
• 临床试验:健康志愿者对SAL200耐受良好,无发热、腹泻等副作用。
三、内溶素在食品安全中的应用
3.1 清除细菌生物被膜
生物被膜是细菌在食品加工设备表面形成的保护性群落,难以清除。
• 金黄色葡萄球菌:LysCSA13、LysK等可清除不锈钢、玻璃表面的生物被膜。
• 单核细胞增生李斯特菌:PlyLM和293-酰胺酶具有抗生物被膜活性。
• 沙门氏菌:Lys68与有机酸联用可协同减少生物被膜。
3.2 在畜牧业中的应用
抗生素在畜牧业中的滥用推动了耐药菌的产生,内溶素成为理想的替代方案:
• 产气荚膜梭菌:phiSM101对家禽病原菌有广谱杀菌作用。
• 沙门氏菌:LysSE24对鸡源沙门氏菌有效。
3.3 在肉类和乳制品中的应用
肉类:LysSA11在4°C下可使火腿中MRSA减少3-log;Trx-SA1可减少奶牛乳房的金黄色葡萄球菌污染。
乳制品:
• LysH5、LysSA97在巴氏奶中对金黄色葡萄球菌有强杀菌效果,LysH5与乳链菌肽有协同作用。
• PlyP100可防止新鲜奶酪中的李斯特菌污染。
• Ctp1L可防止奶酪的“晚期膨胀”缺陷。
3.4 在蔬菜中的应用
• LysP53可裂解生菜上的肠炎沙门氏菌。
• LysWL59、LysWL60可减少生菜上的鼠伤寒沙门氏菌。
• cpp-lys对生菜上的产气荚膜梭菌有效。
3.5 在快速检测中的应用
内溶素的CBD具有高特异性和亲和力,可用于病原菌快速检测:
• 用荧光标记的CBD可特异性识别靶标菌。如CBD-P35和CBD500可在牛奶和奶酪中区分不同李斯特菌株。
• 结合免疫磁分离技术,PlyV12的CBD可高灵敏度检测牛奶中的金黄色葡萄球菌。
• 局限性:CBD难以穿透革兰氏阴性菌的外膜,未来需将其整合到生物传感器技术中。
表1、内溶素在不同食品产品中的应用
| 针对宿主的抗菌活性谱 | 内溶素 | 主要结果 | 食品应用 |
|---|---|---|---|
| S. aureus (金黄色葡萄球菌) | Lysdb | • 破坏 S. aureus 细胞壁中存在的 6-O-乙酰化肽聚糖。 | 奶酪 |
| S. aureus (金黄色葡萄球菌) | LysSA97 | • 在脱脂牛奶和全脂牛奶中与香芹酚(油)具有协同杀菌活性。 | 牛奶 |
| S. aureus (金黄色葡萄球菌) | LysSA11 | • 在 4°C 下 15 分钟内,使实验性污染 MRSA 的肉类中细菌细胞裂解 3-log CFU/mL。 • 钢刀和塑料砧板的去污剂。 | 火腿 |
| S. aureus (金黄色葡萄球菌) | LysH5 | • 牛奶中细菌细胞减少 8 logs CFU/ml。 • 对临床葡萄球菌(包括表皮葡萄球菌和金黄色葡萄球菌)具有广谱宿主范围。 • 在乳制品中与乳酸链球菌素结合使用对 S. aureus 具有协同灭菌活性。 | 牛奶 |
| S. aureus (金黄色葡萄球菌) | HydH5Lyso, HydH5SH3b, CHAPSH3b | • HydH5 和 CHAPSH3b 在 37°C 和 25°C 下对脱脂和全脂牛奶显示出显著的杀菌作用。 | 全脂和脱脂牛奶 |
| S. aureus (金黄色葡萄球菌) | PlyV12 | • 检测牛奶中 S. aureus 的高特异性。 | 加标牛奶 |
| S. aureus (金黄色葡萄球菌) 和 Bacillus cereus (蜡样芽孢杆菌) | LysB4EAD-LysSA11 | • 对 S. aureus 和 B. cereus 具有良好的细胞裂解活性。 • 仅需 2 小时,使用 3.0 μM LysB4EAD-LysSA11 治疗即可将米饭中的 S. aureus 和 B. cereus 降至零水平。 | 米饭 |
| L. monocytogenes (单核细胞增生李斯特菌) | PlyP100 | • 在鲜奶酪 (Queso Fresco) 中可持续 28 天。 • 在奶酪凝块中保留率高。 • 冷藏期间稳定性高。 • 剂量依赖性杀李斯特菌活性。 | 鲜奶酪 (Queso Fresco) |
| L. monocytogenes (单核细胞增生李斯特菌) | Ply511, Ply118 | • 对李斯特菌菌株具有广谱活性。 • 高耐热性。 | 牛奶 |
| L. monocytogenes (单核细胞增生李斯特菌) | PlyP825 | • 与静水压具有协同抗菌活性。 | 牛奶 |
| 单核细胞增生李斯特菌 (L. monocytogenes) | PlyP40 和 PlyPSA | • PlyP40 对多种李斯特菌有效。 • PlyP40 和 PlyPSA 能有效降低新鲜奶酪(queso fresco)中的 L. monocytogenes 数量。 • PlyP40 和 PlyPSA 在冷藏条件下能保持其裂解活性。 | 新鲜奶酪 |
| 单核细胞增生李斯特菌 (L. monocytogenes) | LysZ5 | • 在 4°C 下三小时内,豆浆中的细菌减少 4-log CFU/mL。 | 豆浆 |
| 大肠杆菌 O157:H7 (Escherichia coli O157:H7) | PlyEc2 | • 有效的去污剂。 • 在罗马生菜去污模型中,消除了 99.7% 的 E. coli O157:H7 细胞。 • 处理后的叶片口感和质地不受影响。 | 罗马生菜 |
| 鼠伤寒沙门氏菌 (Salmonella Typhimurium) | LysWL59 和 LysWL60 | • LysWL59 和 LysWL60 对暴露于氯仿的革兰氏阴性菌显示出广泛的催化作用。 • 经 2.5mM LysWL59 和 0.5mM EDTA 处理,生菜上的 S. typhimurium 细胞在 1 小时内减少了 93%。 | 生菜/沙拉 (蔬菜) |
| 产气荚膜梭菌 (Clostridium perfringens) | LysCPAS15 | • 细菌细胞在牛奶中两小时内最多减少 3 logs。 • 快速特异性检测牛奶中的 C. perfringens。 | 牛奶 |
| 丁酸梭菌 (Clostridium tyrobutyricum) | Ctp11 | • 对宿主细菌具有中等的杀菌活性 | 牛奶 |
四、内溶素对主要食源性致病菌的作用
4.1 沙门氏菌
沙门氏菌是常见的食源性病原体,有2000多种血清型。针对鼠伤寒沙门氏菌:
○ LysSTG2、En4 和 SPN9CC 这三种噬菌体来源的内溶素对鼠伤寒沙门氏菌(S. typhimurium)表现出杀菌活性。
○ 具体效果:
• LysSTG2(100 μg/mL)与弱酸性次氯酸水(含氯40 mg/L)联用,可消除超过99%的鼠伤寒沙门氏菌。
• En4 在添加了0.1%碳酸氢钠(NaHCO3)的冷冻和解冻生鸡肉中,使鼠伤寒沙门氏菌减少了1.0–1.6 log CFU/g。
○ 结论: 这些结果表明,将内溶素与增透剂(Permeabilizers)(帮助物质穿透细胞膜的试剂)结合使用,可能是食品工业中对抗鼠伤寒沙门氏菌的一种新颖且有效的方法。
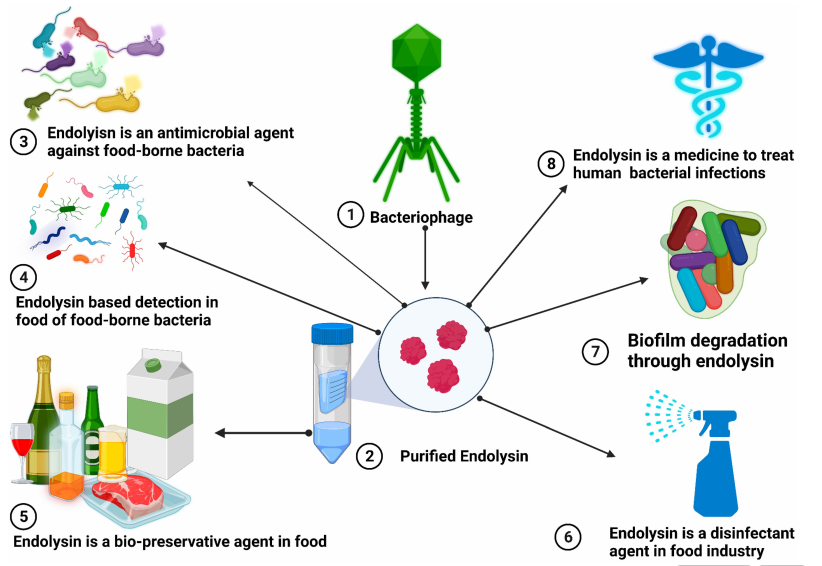
图2、噬菌体衍生内溶素的抗菌特性:在保障人类安全及食品免受致病菌侵害方面的应用
注:内溶素可作为食品链中的生物防腐剂,并用于降解生物膜。此外,内溶素还可用于食源性细菌的食品检测,作为食品工业中的消毒剂,以及在临床中作为治疗人类抗生素耐药性细菌病原体的治疗药物。
4.2 大肠杆菌O157:H7
该菌可导致出血性腹泻甚至死亡,常污染生菜。PlyEc2和RLysJN01能有效杀灭生菜上的该菌而不影响品质。
4.3 金黄色葡萄球菌
MRSA是重要的食源性耐药菌:
• Lys109在牛奶和培根中有效抑制MRSA。
• CHAPSH3b可将牛奶中金黄色葡萄球菌降至检测不到的水平,且耐热耐储存。
4.4 单核细胞增生李斯特菌
该菌危害孕妇、老人等免疫力低下人群:
• PlyP40和PlyPSA可在冷藏28天内清除新鲜奶酪中的李斯特菌。
• PlyP35、Ply511等耐热内溶素在90°C加热30分钟后仍有活性。
• LysZ5在4°C下3小时内使豆奶中活菌减少4-log。
4.5 梭菌属
• 产气荚膜梭菌(Clostridium perfringens):LysCPAS15在牛奶中有效;PlyCP10和PlyCP41抑制猪、鸡源分离株。
• 肉毒梭菌(Clostridium botulinum):CBO1751可裂解该菌并消除神经毒素产生。
• 生孢梭菌(Clostridium sporogenes):CS74L对16株测试菌均有活性。
4.6 副溶血性弧菌
副溶血性弧菌(Vibrio parahaemolyticus)是海产品主要病原体,耐药性问题日益严重(调查显示80%以上对氨苄青霉素耐药)。Lysqdvp001与聚赖氨酸联用对副溶血性弧菌及其生物被膜有协同抑制效果。
4.7 空肠弯曲杆菌
空肠弯曲杆菌(Campylobacter jejuni)是导致全球胃肠道炎最常见的食源性细菌,对公共卫生和食品安全构成严重威胁。
• 内溶素应用: 内溶素 Cj1 和 Cj5 在冷藏(5°C)的鸡肉食品模型中能够根除和控制空肠弯曲杆菌。
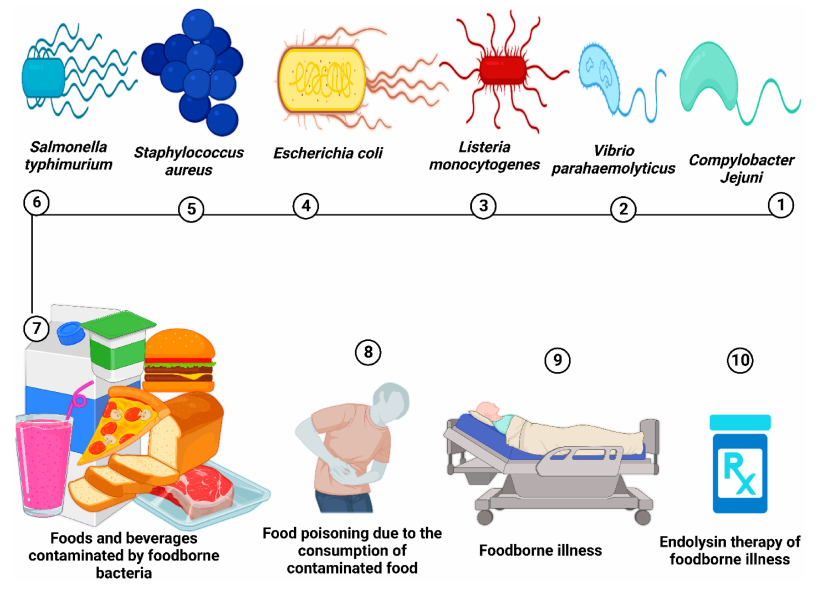
图3、食品受食源性致病菌污染,会导致食物中毒,甚至可能引发严重的食源性疾病。内溶素疗法可作为治疗食源性细菌感染的有效手段之一。
表2、内溶素针对不同病原菌的应用
| Lysin (内溶素) | 抗菌活性谱 | 显著特征 |
|---|---|---|
| PlyC | • 链球菌属 (Streptococcus spp.) | • 一种强效的抗菌内溶素在人体和小鼠模型中进行了测试。 • PlyC 未显示不良反应和过敏反应。 • 作为潜在的安全治疗性内溶素用于治疗细菌感染。 |
| LysAB54 | • 大肠杆菌 (Escherichia coli) • 鲍曼不动杆菌 (Acinetobacter. baumannii) • 铜绿假单胞菌 (Pseudomonas aeruginosa) • 肺炎克雷伯菌 (Klebsiella pneumoniae) | • 天然新型内溶素。 • 快速杀菌活性。 • 100 µg/mL 在 37°C 下孵育 10 分钟可降低 4 个对数级 (4-log)。 • 广谱抗菌活性。 • 对所有 40 种测试的革兰氏阴性菌株均有效。 |
| LysP53 | • 大肠杆菌 (E. coli) • 鲍曼不动杆菌 (A. baumannii) • 铜绿假单胞菌 (P. aeruginosa) • 肺炎克雷伯菌 (K. pneumoniae) | • 工程化嵌合内溶素。 • 经 100 µg/mL 处理 1 小时后,细菌减少 5 个对数级。在小鼠烧伤感染模型中具有较高的去定植功效。 • 广谱抗菌活性。 |
| LysPA26 | • 大肠杆菌 (E. coli) • 肺炎克雷伯菌 (K. pneumonia) • 鲍曼不动杆菌 (A. baumannii) • 铜绿假单胞菌 (P. aeruginosa) | • 天然噬菌体衍生内溶素。 • 50 µg/mL 可在 30 分钟内降低 4 个对数级。 • 消除生物膜形成。 • 保持良好的热稳定性。 |
| PlyE146 | • 大肠杆菌 (E. coli) • 鲍曼不动杆菌 (A. baumannii) • 铜绿假单胞菌 (P. aeruginosa) | • 在 400 µg/ml 剂量下,37°C 处理 2 小时后,阻止了大肠杆菌和铜绿假单胞菌(3 至 3.8 log10 CFU/ml)的生长。 • 鲍曼不动杆菌减少(4.9 至 5 log10 CFU/mL)。 |
| LysABP-01 | • 大肠杆菌 (E. coli) • 鲍曼不动杆菌 (A. baumannii) • 铜绿假单胞菌 (P. aeruginosa) | • 20 µM (500 µg/mL) 抑制细菌生长。 • 破坏大肠杆菌、鲍曼不动杆菌和铜绿假单胞菌菌株的粗糙细胞壁。 |
| Ply6A3 | • 大肠杆菌 (E. coli) • 鲍曼不动杆菌 (A. baumannii) • 铜绿假单胞菌 (P. aeruginosa) • 肺炎克雷伯菌 (K. pneumoniae) • 耐甲氧西林金黄色葡萄球菌 (MRSA) | • Ply6A3 (2 mg/mL) 和溶菌酶 (2 mg/mL) 可降解细菌细胞,并使用扩散法产生透明圈。小鼠腹腔注射后未观察到副作用。 • 内溶素 PD-6A3 和 Ply6A3 的抗菌效果优于 14 种噬菌体的混合物。 |
| LysAB2 | • 鲍曼不动杆菌 (A. baumannii) • 金黄色葡萄球菌 (S. aureus) | • 对多种革兰氏阳性和革兰氏阴性细菌具有很强的抗菌活性。 • 暴露于 500 µg/ml 的 LysAB2 长达 60 分钟后,细菌细胞裂解显著。 • 裂解金黄色葡萄球菌的细胞壁和鲍曼不动杆菌的肽聚糖。 |
| EndoT5 | E. coli (大肠杆菌) | • 在添加透化剂的情况下,对细菌培养稳定期有 4 到 5 log 的裂解活性。 • T5 内溶素与多粘菌素 B (0.4 µg/ml) 或洗必泰 (0.5 µg/mL) 联用可使 CFU 数量减少 5 log,与聚赖氨酸 (80 µg/mL) 联用可减少 4 log。 • 热稳定内溶素;在 90°C 加热 30 分钟后,仍保留 65% 的初始活性。 |
| Lysep3 | • E. coli (大肠杆菌) • P. aeruginosa (铜绿假单胞菌) | • 与 EDTA (25 mM) 联用,Lysep3 可裂解大肠杆菌和铜绿假单胞菌。 • vB_EcoM-ep3 (Lysep3 的噬菌体) 可裂解 15 株来自鸡的多重耐药致病性大肠杆菌临床分离株中的 9 株。 |
| Art-175 | • 大肠埃希菌 (Escherichia coli) 分离株 | • 粘菌素和 Art-175 破坏革兰氏阴性菌的外膜。 • Art-175 杀死了从鸡、牛和猪分离出的所有耐粘菌素大肠杆菌菌株。 • Art-175 与粘菌素之间无交叉耐药性。 |
| AcLys | • E. coli (大肠杆菌) • P. aeruginosa (铜绿假单胞菌) • A. baumannii (鲍曼不动杆菌) • K. pneumoniae (肺炎克雷伯菌) | • 对革兰氏阴性菌具有抗菌活性,导致活菌培养物减少 1.52 (CFU)/mL。 • AcLys 是一种具有 C 末端螺旋结构域的“天然人工化”酶,具有显著的抗革兰氏阴性菌潜力。 |
| LysSS | • E. coli (大肠杆菌) • Salmonella (沙门氏菌) • A. baumannii (鲍曼不动杆菌) • K. pneumoniae (肺炎克雷伯菌) • P. aeruginosa (铜绿假单胞菌) • MRSA (耐甲氧西林金黄色葡萄球菌) | • 革兰氏阳性菌和革兰氏阴性菌均对 LysSS 敏感。 • 显示出抗生物膜活性。 • 对人类细胞无细胞毒性。 • 腹腔注射 LysSS (125 ug/mL) 后,40% 的小鼠从鲍曼不动杆菌全身感染中获救。 |
| Lys68 | • S. typhimurium (鼠伤寒沙门氏菌) • E. coli (大肠杆菌) • P. aeruginosa (铜绿假单胞菌) • P. fluorescens (荧光假单胞菌) • A. baumannii (鲍曼不动杆菌) | • 更广谱的裂解功效(1.5–3 log 降低)。 • 对细菌生物膜及其稳定期具有裂解活性。 • Lys68 对假单胞菌菌株显示出更好的活性。 • 柠檬酸与 Lys68 以及苹果酸与 Lys68 酸具有广泛的抗菌作用,特别是对鼠伤寒沙门氏菌 LT2,导致 3 到 5 log 的减少。 • Lys68 显示出良好的热稳定性。 |
| SPN9CC | E. coli (大肠杆菌) | • 0.5 µg/ml 在 5 分钟内裂解了所有 23 株测试的革兰氏阴性细菌菌株。 • 具有广谱抗菌活性。 • 对数期细菌被杀灭,减少 2 log。 • 使用 OMP (100 mM EDTA) 时,其活性增加,减少量提高四个 log。 • 在 24 度至 65°C 的宽温度范围内具有活性且稳定。 • 在 50°C 时显示出最大裂解活性。 |
| SPN1S | • 鼠伤寒沙门氏菌 (S. typhimurium) • 大肠杆菌 (E. coli) | • 它分为两个亚基,一个用于酶活性,另一个用于肽聚糖结合活性。 • 它对细菌细胞肽聚糖具有良好的裂解活性。 • 对多种革兰氏阴性菌具有杀菌活性。 |
| Ply17 | • 铜绿假单胞菌 (P. aeruginosa) • 大肠杆菌 (E. coli) • 金黄色葡萄球菌 (S. aureus) • 表皮葡萄球菌 (S. epidermidis) | • 测试了不同浓度的Ply17。500 µg/mL的Ply17使经EDTA处理的PAO1活菌数减少了1个对数单位。 • 对对数生长期的细菌具有3个对数级的杀灭活性。 • 在37°C和pH 7.5条件下显示出更好的裂解活性。 • 对革兰氏阳性和革兰氏阴性菌种具有广谱抗菌活性。 • Ply17可以裂解细菌的肽聚糖层。 |
| Lys394 | 大肠杆菌 (E. coli) | • 经过25 µg Lys394、1 mM EDTA和50 µg/ml PGLa肽在室温下处理30分钟后,草坪大肠杆菌菌落形成单位的能力降低了四个数量级。 • 通过计算机模拟氨基酸序列分析,Lys394被鉴定为一种内肽酶。 • 在pH 8.5和低离子强度下,酶活性达到峰值。 • 它显示出以裂解素剂量依赖性方式裂解浮游细菌的活性。 |
| 沙门氏菌噬菌体内溶素 Gp110 (Salmonella phage endolysin Gp110) | • 沙门氏菌 (Salmonella) • 铜绿假单胞菌 (P. aeruginosa) | • 对铜绿假单胞菌PAO1和鼠伤寒沙门氏菌LT2具有抗菌特性。 • 耐热内溶素。 • 在20至90°C温度下,Gp110保持活性。 |
| Ap3gp15 | • 大肠杆菌 (E. coli) • 铜绿假单胞菌 (P. aeruginosa) • 鼠伤寒沙门氏菌 (S. typhimurium) • 肺炎克雷伯菌 (K. pneumoniae) • 伯克霍尔德菌 (B. cenocepacia) | • AP3gp15是一种溶菌酶,可使PG-1,4-糖苷键失活,释放GlcNAc和MurNAc。 • AP3gp15的效力是目前市售溶菌酶的两倍。 • 对肠炎沙门氏菌鼠伤寒亚种、大肠杆菌、肺炎克雷伯菌、铜绿假单胞菌、伯克霍尔德菌和大肠杆菌均有活性。 • 热敏感但在低温下相对稳定。 • 对哺乳动物细胞无细胞毒性作用。 • 50 µg/ml的Ap3gp15对细胞系无副作用。 |
| Ply500 | 单核细胞增生李斯特菌 (L. monocytogenes) | • 对无害李斯特菌 (Listeria innocua) 高度活跃。 • 在10和20 mg/mL酶浓度下处理24小时后,针对10⁵ CFU/mL李斯特菌细胞攻击,分别获得了3个对数级和接近5个对数级的减少。 |
| λSA2-E-Lyso-SH3b 和 λSA2-E-LysK-SH3b | 金黄色葡萄球菌 (S. aureus) | • 在牛奶中活性强烈。 • 3小时内100 µg/ml的浓度可减少加工牛奶中的金黄色葡萄球菌细菌负荷。 |
| ClyC | 金黄色葡萄球菌 (S. aureus) | • 高活性的嵌合溶素。 • 对金黄色葡萄球菌显示出9个对数级的减少。 • 在金黄色葡萄球菌小鼠感染模型中,单次腹腔注射0.1 mg/只的ClyC显著提高了存活率。 • 它使受感染小鼠器官中的细菌负荷降低了2个对数级 (CFU/mL)。 |
| ClyH | 金黄色葡萄球菌 (S. aureus) | • 对浮游MRSA细胞具有活性。 • ClyH通过细胞裂解活性以时间依赖性方式根除MRSA生物膜。 • 活菌平板计数和动力学分析显示,不同菌龄的生物膜均对ClyH敏感。 • 10 µg/ml的ClyH减少了甲氧西林敏感性金黄色葡萄球菌 (MSSA) 和MRSA菌株的生物膜生物量。 |
| ClyH | 金黄色葡萄球菌 (S. aureus) | • ClyH对金黄色葡萄球菌菌株的最小抑菌浓度 (MIC) 范围为0.05至1.61 mg/L。 • 在小鼠感染模型中,一剂ClyH保护小鼠免受MRSA感染导致的死亡。 • 无副作用。没有副作用的证据。 |
| ClyF | MRSA (耐甲氧西林金黄色葡萄球菌) | • ClyF具有最佳的MRSA生物膜破坏活性。 • 对广泛的金黄色葡萄球菌菌株具有抗菌活性。 • 在小鼠菌血症和伤口感染模型中,单次ClyF治疗显示出良好的MRSA清除活性。 |
| StBP1-ClyF | 金黄色葡萄球菌 (S. aureus) | • 对金黄色葡萄球菌具有良好的杀菌活性。 • 强效的抗菌和抗生物膜特性。 • 12.5 µg/mL的蛋白质,触发活菌数量下降5.52个对数级。 • 有效杀灭MRSA(1小时内>99.999%)。 • 抑制各种表面上动态和静态的金黄色葡萄球菌生物膜生长,包括硅胶导管、硅化玻璃和硅胶涂层乳胶导管。 • 在固体支撑表面上具有有效的固定化可靠性。 |
| LyS15S6 | 沙门氏菌 (Salmonella) | • 高酶活性。 • 针对大肠杆菌、志贺氏菌、铜绿假单胞菌、鲍曼不动杆菌和肺炎克雷伯菌的广谱裂解活性。 • 良好的热稳定性。 • 对沙门氏菌具有强杀菌活性。 • 1 µg/mL EPL (ε-聚-L-赖氨酸),2 µM LyS15S6可以在25°C下孵育2小时后,使测试的肠杆菌科细菌活细胞减少3-4个对数级。 • 在25°C孵育15分钟和8°C孵育2小时后,沙门氏菌分别减少了2.56和3.14个对数级。 |
| LysPBC1 | 蜡样芽孢杆菌 (Bacillus cereus) | • 快速杀死蜡样芽孢杆菌宿主细菌。 • 宿主特异性广。 • 针对蜡样芽孢杆菌的生物防治剂。 • 裂解所有蜡样芽孢杆菌群细菌,包括蜡样芽孢杆菌 (B. cereus)、伟氏蜡样芽孢杆菌 (B. weihenstephanensis)、蕈状芽孢杆菌 (B. mycoides)、伟氏蜡样芽孢杆菌 (B. weihenstephanensis) 和苏云金芽孢杆菌 (B. thuringiensis)。 |
| Psa 和 Psm | 产气荚膜梭菌 (C. perfringens) | • Psa 对产气荚膜梭菌具有高裂解杀菌活性。 • 1.25 µg/mL Psa 和 3.9 ± 0.16 µg/mL Psm 具有良好的协同活性。 • Psa 和 Psm 联合使用有助于治疗和预防产气荚膜梭菌感染。 |
| LysCPS2 | 产气荚膜梭菌 (C. perfringens) | • 广谱抗菌活性。 • 对产气荚膜梭菌菌株具有高度特异性。 • 高热稳定性内溶素。 • 在 95°C 反应 10 分钟后,仍保留 30% 针对产气荚膜梭菌的催化活性。 • 在 pH 7.5–10 和温度 25–65°C 下杀菌活性最佳。 • 在宽范围的 NaCl 浓度下高度稳定。 • 针对产气荚膜梭菌的检测和生物防治剂。 |
| LysSE24 | 多重耐药沙门氏菌菌株 | • 对 23 种多重耐药沙门氏菌菌株具有广谱活性。 • LysSE24 在 pH 4.0 至 10.0 和温度 20 至 60°C 下相对稳定。 • 0.1 µM LysSE24 反应长达 5 分钟可使肠炎沙门氏菌 (Salmonella Enteritidis) 变性。 |
| LysSP1 | 鼠伤寒沙门氏菌 (S. typhimurium) | • 对革兰氏阳性和革兰氏阴性食源性细菌病原体均具有良好的杀菌活性。 • 10 µg LysSP1 与 EDTA 结合可杀死所有 10⁶ CFU/mL 的沙门氏菌细胞。 • 对沙门氏菌菌株、大肠杆菌 (E. coli)、大肠杆菌 O157、金黄色葡萄球菌 (S. aureus)、单核细胞增生李斯特菌 (L. monocytogenes) 和志贺氏菌 (Shigella) 显示出裂解活性。 • 在 40°C 时活性最佳。 • 在 4°C 下稳定 7 天,在 -20°C 下稳定 180 天。 • 对革兰氏阴性和革兰氏阳性细菌菌株均有效。 |
| LysSTG2 | 鼠伤寒沙门氏菌 (S. typhimurium) | • 高热稳定性。 • 在 50°C 加热后仍保持 93% 的裂解活性。 • 100 µg/mL 针对鼠伤寒沙门氏菌 NBRC 12529 浮游细胞及其生物膜,在孵育 1 小时后显示出 1.2 log 的减少。 • 40 mg/L 的氯和 100 µg/mL 的 LysSTG2 消除了超过 99% 的生物膜细胞。 |
| LysT144 | 鼠伤寒沙门氏菌 (S. typhimurium) | • 针对沙门氏菌的广谱活性。 • 2 µg/mL 对鼠伤寒沙门氏菌具有快速且显著的裂解活性。 • 在 30 分钟反应时间内,将经氯仿处理的鼠伤寒沙门氏菌的 OD600nm 从 0.80 降低到 0.14。 |
五、当前面临的挑战
5.1 稳定性问题
食品成分(盐、酸、蛋白、脂肪等)及温度、pH等因素可能影响内溶素的稳定性和活性。如LysAB54在牛奶中活性丧失。未来可通过封装(如纳米囊泡)和计算辅助设计来提高稳定性。
5.2 对静止期细菌活性低
细菌在静止期细胞壁交联更紧密,内溶素效果下降。可通过与其他抗菌剂联用(EDTA、柠檬酸、壳聚糖)或组合不同内溶素来增强协同效应。
5.3 革兰氏阴性菌的外膜屏障
这是内溶素应用的核心障碍。主要解决策略包括:
表3、革兰氏阴性菌外膜屏障的解决策略及原理
| 策略 | 原理 |
|---|---|
| 物理/化学处理 | 破坏外膜完整性,如栅栏技术 |
| 融合通透性肽 | 融合天蚕素A等阳离子肽 |
| 与抗生素联用 | 协同增效(如LNT113) |
| 工程化改造 | 开发人工溶素、溶素、内溶素等 |
| 封装递送 | 使用脂质体、壳聚糖纳米颗粒等 |
5.4 生产成本高
内溶素的规模化生产、纯化成本较高,限制了食品工业的广泛应用。需在安全性和稳定性评估基础上,优化生产工艺以降低成本。
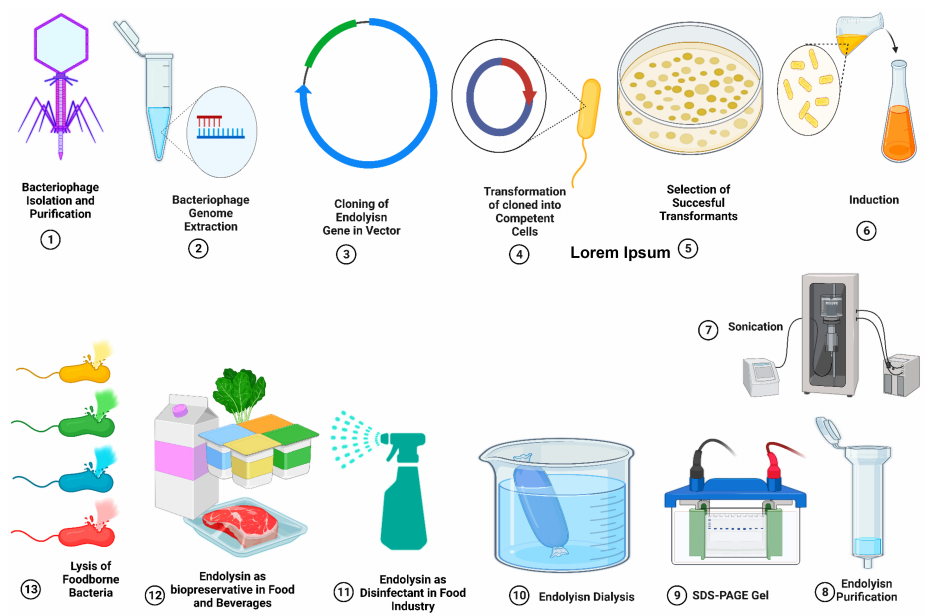
图4、噬菌体衍生内溶素的克隆、表达和纯化。内溶素作为天然生物防腐剂的应用,旨在减少食源性致病菌对食品的污染。
六、内溶素的工程化改造
内溶素的模块化结构为其工程化改造提供了便利。主要改造策略如下:
6.1 结构域截短或缺失
去除特定结构域可改变内溶素的活性、特异性和结合能力。例如:
• CHAPk(LysK的截短变体)活性优于天然酶。
• 去除CBD后,催化结构域的正电荷增加,增强了与带负电的脂磷壁酸的相互作用,从而提升活性。
6.2 定点诱变
通过改变关键氨基酸提高活性和热稳定性:
突变CD27L催化位点附近的氢键网络残基,提升了对多种李斯特菌的裂解活性。
• 改变Cpl-7S中CBD的15个氨基酸,增加正电荷,提高活性。
• 改造LysF1的疏水核心,提高热稳定性。
6.3 结构域重组(嵌合酶)
将不同内溶素的结构域进行重组,可创造兼具多重优势的嵌合酶:
• CHAPSH3b(HydH5的CHAP结构域+溶葡萄球菌素的SH3b结构域):在巴氏奶中15分钟内使金黄色葡萄球菌降至无法检测水平。
• ClyR(通过高通量筛选获得):对停乳链球菌有广谱裂解活性。
• ClyC(重排12种葡萄球菌内溶素结构域):在牛奶和动物血液中活性稳定,并能保护小鼠免受MRSA感染。
6.4 针对革兰氏阴性菌的改造
• 人工溶素(Artilysins):内溶素融合外膜去稳定肽(如Art-175)。
• 溶素(Lysocins):内溶素与细菌素融合,利用细菌素穿透外膜。
• 内溶素(Innolysins):内溶素融合噬菌体受体结合蛋白(如Innolysin Ec21)。
• 封装技术:使用脂质体或壳聚糖纳米颗粒包埋内溶素,提高其穿透性和稳定性。
七、结论与展望
内溶素作为一种新型抗菌蛋白,在食品安全领域展现了广阔的应用前景。目前其主要应用于革兰氏阳性菌防控,对革兰氏阴性菌的效力仍需进一步提升。未来的研究应聚焦于:
✅ 针对性设计:针对特定食品基质中的特定病原体,开发定制化内溶素。
✅ 优化性能:兼顾高活性、热稳定性、溶解性和成本效益。
✅ 拓展应用:将工程化内溶素从临床推广到食品工业场景。
随着分子生物学工具的不断进步,工程化内溶素有望成为控制食源性致病菌的理想工具,为食品安全提供更可靠的技术保障。
参考文献
1.AbhisinghaM.DumnilJ.PitaksutheepongC. (2023). Effect of lysin EN4 in combination with sodium bicarbonate on reduction of Salmonella in chilled and thawed chicken meat. Int. J. Food Microbiol.387:110058. doi: 10.1016/j.ijfoodmicro.2022.110058
2.GondilV. S.KhanF. M.MehraN.KumarD.KhullarA.SharmaT.et al. (2021). “Clinical potential of bacteriophage and Endolysin based therapeutics: A futuristic approach” in Microbial Products for Health, Environment and Agriculture. ed. AroraP. K. (London: Springer Nature), 39–58.
3.Li C., Nyaruaba R., Zhao X., Xue H., Yang H., Li Y., et al. (2023). LysP53 activity against Salmonella and its application in decontamination of Salmonella on fresh romaine lettuce. Appl. Microbiol. Biotechnol.107, 5403–5413. doi: 10.1007/s00253-023-12666-2
4.Shen K.-S., Shu M., Tang M.-X., Yang W.-Y., Wang S.-C., Zhong C., et al. (2022). Molecular cloning, expression and characterization of a bacteriophage JN01 endolysin and its antibacterial activity against E. coli O157: H7. LWT165:113705. doi: 10.1016/j.lwt.2022.113705
5.Stone E., Pennone V., Reilly K., Grant I. R., Campbell K., Altermann E., et al. (2022). Inhibition of Listeria monocytogenes by phage lytic enzymes displayed on tailored bionanoparticles. Foods11:854. doi: 10.3390/foods11060854
相关产品
HZB124202:产气荚膜梭菌 | Clostridium perfringens
HZB353719:鲍曼不动杆菌 | Acinetobacter baumannii
HZB364558:大肠埃希氏菌 | Escherichia coli
HZB364135:铜绿假单胞菌 | Pseudomonas aeruginosa
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-06-30
编制人:思琪
审稿人:叶凡