质粒入门指南2:质粒是什么?(下)(Whatisa Plasmid?)(2/2)
来源:武汉市灰藻生物科技有限公司 浏览量:13 发布时间:2026-06-23 17:59:52
四、启动子区域
既然我们已经能够复制质粒并确保细胞维持其存在,下一步就是让质粒表达我们的目的基因。这就引出了启动子——负责启动插入片段转录的元件。
在实际应用中,“启动子”一词通常指代启动子(RNA 聚合酶结合位点)与操纵基因(响应元件)的组合。启动子的长度约为 100 到 1000 个碱基对,通常位于其靶基因的上游。启动子区域的序列控制着 RNA 聚合酶和转录因子的结合,因此启动子在决定目的基因的表达位置和时间方面发挥着重要作用。
1. RNA 聚合酶
RNA 是通过 RNA 聚合酶(RNAP)从 DNA 转录而来的。在细菌中,这一过程由单一酶完成;而在真核生物中,存在多种聚合酶,每种聚合酶负责转录特定类别的 RNA。为了实现这种特异性,真核生物的 RNA 聚合酶能够识别并结合特定的启动子元件。这意味着您质粒骨架中存在的启动子必须与所需合成的 RNA 类型相匹配:如果您需要信使 RNA(mRNA,用于基因表达),则需要使用 RNA 聚合酶 II 启动子;而小分子 RNA(如短发夹 RNA,shRNA)则由 RNA 聚合酶 III 启动子转录。
本节主要介绍用于一般 RNA 聚合酶 II 和 RNA 聚合酶 III 转录的启动子;不过,在慢病毒和逆转录病毒构建体中,也常采用如 RNA 聚合酶 II 启动子等病毒长末端重复序列(LTR)。这些内容将在我们的病毒载体资源中另行介绍。
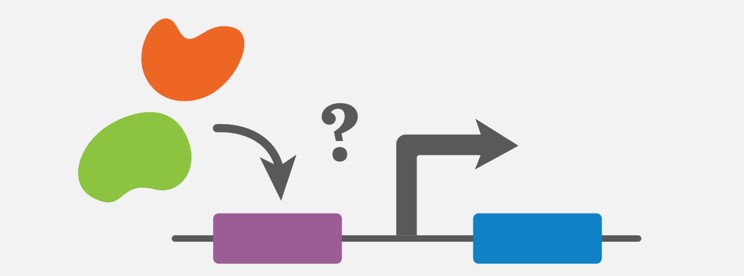
图 3:转录因子与基因(蓝色)的启动子区域(紫色)结合
2. 启动子的特异性
除了根据 RNA 转录本的类型选择启动子外,您还需要确保质粒上的启动子适合在您的宿主生物体中发挥作用。由于不同细胞类型或生物体之间的转录机制存在差异,启动子也必须具有相应的多样性。细菌启动子仅在原核细胞中起作用,且通常仅在其来源物种或亲缘关系相近的物种中起作用。
同样,各种真核细胞类型(如哺乳动物细胞、酵母、植物等)也需要独特的启动子,且它们之间几乎没有交叉通用性。一般来说,细菌启动子的多样性和复杂性较低,其组成部分少于真核细胞启动子。有些启动子具有组成型活性,始终处于开启状态;而另一些启动子则受到更为严格的调控。
受调控的启动子可能仅在特定的组织中或发育的特定阶段发挥作用,或者可以通过化学物质、温度或光照等外部条件按需开启或关闭。在细胞内,启动子本身还受到其他调控因子的控制,如增强子、边界元件、绝缘子和沉默子;然而,仍可能会发生一定程度的“渗漏”转录。通常情况下,这对细胞来说不是大问题,但如果您的目的基因具有毒性,渗漏转录可能会干扰研究结果,甚至导致细胞死亡。
为了解决这一问题,科学家们构建了人工合成启动子。这些启动子通常包含多种其他启动子元件的组合,往往受到更严密的调控。
3. 真核生物与原核生物的常见启动子
表 1.3 和表 1.4 列出了一些最常见的细菌和哺乳动物启动子。这些列表绝非详尽无遗,但在您试图挑选最合适的启动子时,可以作为很好的起点。
表 1.3 - 真核生物启动子
| 启动子 | 主要用途 | RNA 转录本 | 描述 | 表达类型 | 其他注意事项 |
|---|---|---|---|---|---|
| CMV | 通用表达 | mRNA | 四环素响应元件启动子 | 可通过四环素或其衍生物诱导 | 通常包含一个基础活性极低的最小化启动子和几个四环素操纵基因。根据所使用的四环素反式激活因子,转录可被开启或关闭。 |
| UAS | 通用表达 | mRNA | 含有 Gal4 结合位点的果蝇启动子 | 特异性表达 | 需要存在 Gal4 基因才能激活该启动子。 |
| Ac5 | 通用表达 | mRNA | 源自果蝇肌动蛋白 5c (Actin 5c) 基因的强效昆虫启动子 | 组成型表达 | 常用于果蝇表达系统。 |
| Polyhedrin | 通用表达 | mRNA | 源自杆状病毒的强效昆虫启动子 | 组成型表达 | 常用于昆虫细胞表达系统。 |
| CaMKIIa | 通用表达 | mRNA | 钙/钙调蛋白依赖性蛋白激酶 II 启动子 | 特异性表达 | 用于神经元/中枢神经系统 (CNS) 的表达。受钙离子和钙调蛋白的调节。 |
| GAL1, 10 | 通用表达 | mRNA | 酵母中相邻且反向转录的启动子 | 半乳糖诱导;葡萄糖阻遏 | 可单独或共同使用。受 GAL4 和 GAL80 调控。 |
| TEF1 | 通用表达 | mRNA | 酵母转录延伸因子启动子 | 组成型表达 | 类似于哺乳动物的 EF1α 启动子。 |
| GDS | 通用表达 | mRNA | 源自酵母 3-磷酸甘油醛脱氢酶的强效启动子 | 特异性表达 | 表达量极高,也称为 TDH3 或 GAPDH。 |
| ADH1 | 通用表达 | mRNA | 酵母乙醇脱氢酶 I 启动子 | 受乙醇阻遏 | 全长版本表达量高且表达强烈;截短版本为组成型表达,但表达量较低。 |
| CaMV35S | 通用表达 | mRNA | 源自花椰菜花叶病毒的强效植物启动子 | 组成型表达 | 在双子叶植物中具有活性,在单子叶植物中活性较低,在动物细胞中具有一定活性。 |
| Ubi | 通用表达 | mRNA | 源自玉米泛素基因的植物启动子 | 组成型表达 | 在植物中能够实现高水平的基因表达。 |
| H1 | 小分子 RNA 表达 | shRNA | 源自人类 RNA 聚合酶 III 启动子 | 组成型表达 | 表达水平可能略低于 U6 启动子,但在神经细胞中可能具有更好的表达效果。 |
| U6 | 小分子 RNA 表达 | shRNA | 源自人类 U6 小核启动子 | 组成型表达 | 也可使用小鼠 U6 启动子,但其效率可能较低。 |
表 1.4 - 原核生物启动子
| 启动子 | 主要用途 | RNA 转录本 | 描述 | 表达模式 |
|---|---|---|---|---|
| T7 | 体外转录 / 通用表达 | 源自 T7 噬菌体的启动子 | 组成型表达,但需要 T7 RNA 聚合酶。 | 仅当基因两侧存在两个方向相反的噬菌体启动子时,才可用于体外转录。 |
| T7lac | 高水平基因表达 | 源自 T7 噬菌体的启动子,附加 lac 操纵基因 | 未诱导时基础表达量极低。需要 T7 RNA 聚合酶(该酶同样受 lac 操纵基因控制)。可被 IPTG 诱导。 | 常见于 pET 载体中。受 lac 操纵基因极其严密的调控。非常适合通过调节诱导剂浓度来调控基因表达。 |
| Sp6 | 体外转录 / 通用表达 | 源自 Sp6 噬菌体的启动子 | 组成型表达,但需要 SP6 RNA 聚合酶。 | 仅当基因两侧存在两个方向相反的噬菌体启动子时,才可用于体外转录。 |
| trp | 高水平基因表达 | 源自大肠杆菌色氨酸操纵子的启动子 | 可阻遏型 | 当细胞内色氨酸水平较高时,该启动子会被关闭。 |
| lac | 通用表达 | 源自 lac 操纵子的启动子 | 在缺乏 lac 阻遏蛋白(lacI 或 lacIq)的情况下呈组成型表达。可被 IPTG 或乳糖诱导。 | 属于渗漏型启动子,表达量相对较弱。lacIq 突变可使阻遏蛋白的表达量增加 10 倍,从而加强对 lac 启动子的调控。非常适合通过调节诱导剂浓度来调控基因表达。 |
| Ptac | 通用表达 | lac 和 trp 的杂合启动子 | 调控方式类似于 lac 启动子 | 包含 trpB 的 -35 区和 lac 的 -10 区。调控极其严密。非常适合通过调节诱导剂浓度来调控基因表达。通常其表达量优于单独的 lac 启动子。 |
| pL | 高水平基因表达 | 源自 λ 噬菌体的启动子 | 可进行温度调控 | 通常与温度敏感型 cI857 阻遏蛋白配对使用。 |
| araBAD | 通用表达 | 阿拉伯糖代谢操纵子的启动子 | 可被阿拉伯糖诱导;在葡萄糖存在时通过分解代谢阻遏作用被抑制,或通过抗诱导剂岩藻糖的竞争性结合被抑制 | 表达量较弱。常见于 pBAD 载体中。非常适合快速调控且基础表达量极低;但不太适合通过调节诱导剂浓度来精细调控基因表达。 |
五、终止子与多聚腺苷酸化信号
根据所包含的功能元件,旨在特定宿主细胞类型中表达基因的质粒通常可分为原核生物和真核生物两大类。在原核和真核系统中,质粒 DNA 都必须被转录为 RNA。转录过程分为三个阶段:起始、延伸和终止。我们在前文讨论了启动子在基因转录起始步骤中的作用;在此,我们将概述转录是如何停止的,即终止(Termination)过程。
1. 什么是终止与多聚腺苷酸化?
终止子是一种基于序列的元件,其作用是界定转录单元(如基因)的末端,并启动将新合成的 RNA 从转录机制上释放的过程。终止子位于待转录基因的下游,通常紧接在任何 3' 端调控元件(如多聚腺苷酸化或多聚 A 信号)之后。虽然许多研究将启动子强度视为决定基因表达水平的因素,但终止子在 RNA 加工中也发挥着重要作用,它会影响 RNA 的半衰期,并最终影响基因的表达水平。
顾名思义,多聚腺苷酸化(Polyadenylation)是指在转录后向信使 RNA 转录本的尾部添加多个腺嘌呤(A)核苷酸的过程。多聚腺苷酸化的目的和机制在不同细胞类型中有所不同,但总体而言,它在真核生物中有助于延长转录本的寿命,而在原核生物中则促进转录本的降解。
2. 原核生物的终止
原核生物的终止机制主要分为两类:依赖 Rho 因子(Rho-dependent)和不依赖 Rho 因子(Rho-independent)。Rho 因子是一种解旋酶,协助 RNA 聚合酶终止转录。基于质粒的表达系统通常不使用依赖 Rho 因子的终止子,因此此处不作详述(文末提供了相关参考文献)。
几乎所有常见的细菌表达质粒都使用不依赖 Rho 因子的终止子,包括天然存在的终止子(如 T7 和 rrnB)以及工程化的高效终止子(如 T0)。不依赖 Rho 因子的终止也被称为内在终止(Intrinsic termination),它依赖于 RNA 转录本中富含 G-C 的发夹结构的形成,随后是一个结合力较弱的多聚尿嘧啶(Poly-U)序列(如右图所示)。发夹-DNA 复合物的三级结构被认为会破坏转录复合物的稳定性,从而启动转录本的切割。
没有任何一种终止子能以 100% 的效率停止模板的转录并启动预期的切割事件,尽管一些工程化终止子的效率已非常接近(>95%)。然而,对于大多数应用而言,任何常见的终止子都足以胜任。许多商业表达载体使用双终止子来减少下游元件的意外翻译。在多顺反子构建体中,为了将转录通读降至最低,可能需要使用高亲和力的终止子。Chris Voigt 实验室对一系列原核生物终止子进行了表征,并将其中几种保存在 Addgene 中。
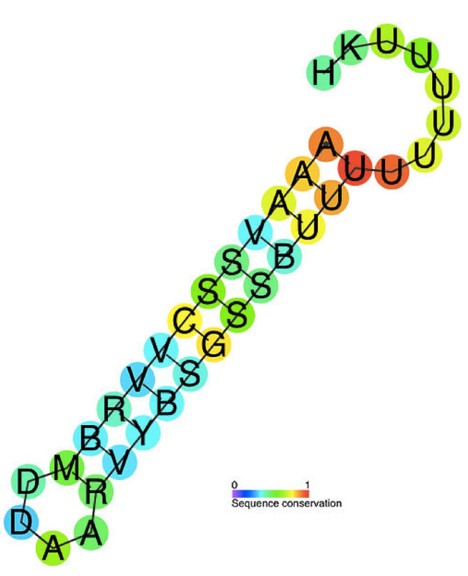
图 4:90个细菌元件的预测保守二级结构和序列保守Rho独立终止注释
3. 原核生物的多聚腺苷酸化
尽管多聚腺苷酸化通常被认为是真核生物特有的过程,但原核生物也会向某些 RNA 添加多聚 A 尾。与真核生物需要共有序列才能添加多聚 A 尾的机制不同,原核生物转录本的多聚 A 尾添加是非特异性的,可以添加到任何可及的 3' 端。多聚 A 尾的存在会将 RNA 靶向至降解体(Degradosome),该复合体包含能够切割未受二级结构保护的 RNA 的酶。由于缺乏特异性,人们认为多聚 A 尾被用于控制调节性 RNA 的细胞内浓度,并且可能还作为一种质量控制机制,帮助细胞清除错误折叠的 RNA。
4. 真核生物的终止与多聚腺苷酸化
与仅有一种 RNA 聚合酶用于转录的原核生物不同,真核生物拥有三种 RNA 聚合酶(聚合酶 I、II 和 III),每种负责转录不同类型的 RNA:聚合酶 I 负责核糖体 RNA(rRNA),聚合酶 II 负责信使 RNA(mRNA)和微小 RNA(miRNA),聚合酶 III 负责转运 RNA(tRNA)和其他短链 RNA。尽管对真核生物终止机制的研究不如原核生物深入,但其基本过程已被阐明,且每种真核生物 RNA 聚合酶的终止方式各不相同。例如,聚合酶 III 依赖于特定的序列和 RNA 二级结构来诱导转录本切割,这与原核生物中的不依赖 Rho 因子的终止类似。这与聚合酶 I 和 II 不同,后两者都需要终止因子的结合。
尽管聚合酶 I 和 II 都依赖终止因子,但它们采用了不同的机制来终止转录。聚合酶 I 使用类似于原核生物依赖 Rho 因子的机制;而聚合酶 II 的终止更为复杂,涉及两种 RNA 聚合酶相关蛋白(CPSF 和 CstF),它们负责招募切割和多聚腺苷酸化酶,这一过程似乎将终止与多聚腺苷酸化偶联在一起。
哺乳动物表达质粒主要用于生成 mRNA。常用的哺乳动物终止子(SV40、hGH、BGH 和 rbGlob)包含 AAUAAA 序列基序,该基序可同时促进多聚腺苷酸化和转录终止。在上述终止子中,由于存在额外的辅助序列,SV40 晚期多聚 A 和 rbGlob 多聚 A 被认为在终止转录方面更为高效(Schek et al., 1992; Gil et al., 1987)。
如前所述,聚合酶 II 转录本(即 mRNA)的终止和多聚腺苷酸化是相互协调的过程。在共有序列基序与下游富含 G-U 的区域之间进行切割(如下图所示),将 mRNA 从聚合酶上释放,并产生一个游离的 3' 端,此时该末端即可进行多聚腺苷酸化。多聚 A 尾的添加对于 mRNA 的稳定性、防止降解至关重要,并且对核输出和翻译过程也起着不可或缺的作用。
六、DNA 甲基化与限制性内切酶
您是否曾在使用 XbaI 或 ClaI 等限制性内切酶进行酶切时,得到过异常或出乎意料的结果?或者您是否思考过,为什么 DpnI 能够降解聚合酶链式反应(PCR)中的模板 DNA,却不会破坏定点突变实验中新生成的产物?这两个问题的答案都指向同一个因素——DNA 甲基化!请继续阅读,了解 DNA 甲基化是如何影响限制性内切酶酶切实验的。
1. 为什么 DNA 会发生甲基化?
除了限制-修饰系统外,DNA 甲基化在调控基因组复制、修复 DNA 合成过程中发生的错配碱基对或微小插入/缺失(Indels),以及促进或抑制蛋白质表达等方面也发挥着不可或缺的作用。限制性内切酶实际上是原核生物用来抵御外源 DNA 的天然限制-修饰系统的一半。该系统的另一半是甲基转移酶,它们会在特定的 DNA 序列上添加甲基,以防止这些序列被限制性内切酶降解。通常,特定的原核生物含有一种或几种限制-修饰系统的编码基因,这些系统包含向特定 DNA 序列添加甲基的甲基转移酶,以及能够识别并在序列未被甲基化时(例如该序列来自噬菌体)对其进行切割的配套限制性内切酶。
参与上述调控和修复过程的甲基化酶(例如 Dam 甲基化酶和 Dcm 甲基化酶)独立于限制-修饰系统,但它们依然会影响某些限制性内切酶能否有效切割 DNA。
常见的实验室大肠杆菌 K12 菌株(如 DH5α)含有 3 种甲基化酶,它们分别识别并甲基化不同的 DNA 序列:
o Dam 甲基化酶:向 DNA 中 GATC 序列的腺嘌呤(A)添加甲基。
o Dcm 甲基化酶:向 CCWGG 序列的第二个胞嘧啶(C)添加甲基。
o EcoKI 甲基化酶:向 AACNNNNNNGTGC 或 GCACNNNNNNGTT 序列中的腺嘌呤添加甲基。
2. 对克隆和酶切实验的影响
尽管并非所有原核生物的 DNA 甲基化程度都相同,但在进行 DNA 酶切时仍应考虑甲基化的潜在影响。为什么呢?因为尽管 Dam 甲基化位点并不专门与任何限制-修饰系统相关联,但其序列可能与限制性内切酶的识别位点重叠,从而抑制 ClaI 或 XbaI 等酶的活性;相反,这也会激活如 DpnI 等酶的活性。
例如,如果 XbaI 的识别位点(TCTAGA)前面紧接 GA 或后面紧跟 TC,其切割作用可能会因甲基化而受阻。如下图所示,由于该限制性内切酶识别位点后紧跟 TC,Dam 甲基化酶的识别位点(红色下划线部分)与 XbaI 的切割位点(橙色线表示)发生了重叠。在该位点添加甲基会阻止酶在此处切割 DNA,不过其他的限制性内切酶识别位点(包括前面没有 GA 或后面没有 TC 的其他 XbaI 位点)仍会被正常切割。如果不将这一因素考虑在内,这种被阻断的切割位点可能会导致实验结果难以解释,甚至让您误以为质粒构建不正确!
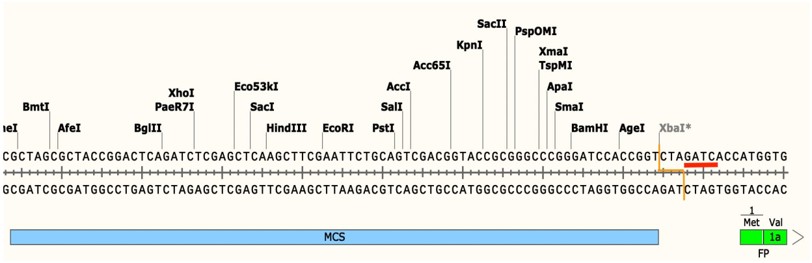
图 5:DNA甲基化对克隆和酶切实验的影响
相反,像 DpnI 这样的酶需要在其识别位点发生甲基化才能有效切割 DNA。DpnI 常用于定点突变实验。在此过程中,通过 PCR 将目标突变引入目的质粒时,生成的突变质粒是不含甲基的(因为 PCR 反应体系中不存在甲基转移酶)。另一方面,模板质粒应来源于 Dam 阳性(dam+)的大肠杆菌菌株,因此其质粒中所有 GATC 序列的腺嘌呤都是甲基化的。当使用 DpnI 消化 PCR 产物时,只有未突变且甲基化的模板会被破坏,从而留下突变质粒池,后续可通过桑格测序(Sanger sequencing)进行验证。
3. 如何判断我的酶是否会被甲基化影响?
要确定您的限制性内切酶是否受甲基化影响,您可以查阅酶供应商提供的说明书或在线数据库,确认该酶的识别位点是否与已知的甲基化位点(如 Dam 或 Dcm 位点)重叠。如果存在重叠,且您的 DNA 来源于具有相应甲基化酶活性的菌株,那么该酶切位点很可能会受到阻断或需要甲基化才能切割。
4. 如何控制甲基化
无论是为了克隆还是诊断目的进行酶切,我们都建议您仔细核对,确保实验结果不会受到甲基化的干扰。幸运的是,实验室中常用的大多数限制性内切酶的识别位点都不会与甲基化位点重叠。下方的快速参考表列出了 10 种可能受甲基化影响的常见酶。该表并非详尽无遗,如需获取更详细的信息,建议您查阅 REBASE 数据库。
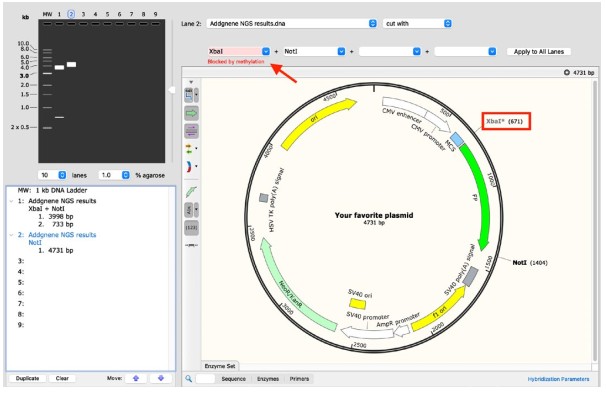
图 6:使用Snapgene(R)创建的图像
最后,您可以通过改变所使用的细菌菌株来控制甲基化。例如,如果您必须使用会被 Dam 或 Dcm 甲基化阻断的限制性内切酶位点,您可以先将 DNA 克隆到 Dam/Dcm 甲基化酶缺陷型大肠杆菌菌株(如 JM110)中,然后重新纯化质粒。这些专门构建的大肠杆菌菌株被设计为缺乏 Dam 和 Dcm 甲基化酶活性,因此它们产生的 DNA 在这些位点上是未甲基化的。请注意,dam-/dcm- 菌株的突变率可能较高(因为它们同样缺乏 Dam 介导的错配修复功能),因此这些菌株不应被用于质粒的长期保存。
表 1.5 - 受甲基化影响的限制性内切酶
| 限制性内切酶 | Dam 甲基化的影响 | Dcm 甲基化的影响 | EcoKI 甲基化的影响 |
|---|---|---|---|
| ApaI | 不受影响 | 因位点重叠而受阻 | 不受影响 |
| BsaI | 不受影响 | 因位点重叠而受阻 | 不受影响 |
| ClaI | 因位点重叠而受阻 | 不受影响 | 不受影响 |
| DraI | 不受影响 | 不受影响 | 因位点重叠而受阻 |
| HpaI | 不受影响 | 不受影响 | 因位点重叠而受阻 |
| MboI | 因位点重叠而受阻 | 不受影响 | 不受影响 |
参考文献与相关资料
https://www.addgene.org/
Peters JM, Vangeloff AD, Landick R. Bacterial Transcription Terminators: The RNA 3′-End Chronicles. Journal of molecular biology. 2011. PubMed PMID: 21439297. PubMed Central PMCID: PMC3622210.
Schek N, Cooke C, Alwine JC. Definition of the upstream efficiency element of the simian virus 40 late polyadenylation signal by using in vitro analyses. Molecular and Cellular Biology. 1992. PubMed PMID: 1333042. PubMed Central PMCID: PMC360476.
Gil A, Proudfoot NJ. Position-dependent sequence elements downstream of AAUAAA are required for efficient rabbit beta-globin mRNA 3’ end formation. Cell. 1987. PubMed PMID: 3568131.
Hager S, Frame FM, Collins AT, Burns JE, Maitland NJ. An Internal Polyadenylation Signal Substantially Increases Expression Levels of Lentivirus-Delivered Transgenes but Has the Potential to Reduce Viral Titer in a Promoter-Dependent Manner. Hum Gene Ther. 2008. PubMed PMID: 18627247.
Zufferey R, Donello JE, Trono D, Hope TJ. Woodchuck Hepatitis Virus Posttranscriptional Regulatory Element Enhances Expression of Transgenes Delivered by Retroviral Vectors. Journal of Virology. 1999. PubMed PMID: 10074136. PubMed Central PMCID: PMC104046.
Wodrich H, Schambach A, Kräusslich H-G. Multiple copies of the Mason–Pfizer monkey virus constitutive RNA transport element lead to enhanced HIV-1 Gag expression in a contextdependent manner. Nucleic Acids Research. 2000. PubMed PMID:10648781. PubMed Central PMCID: PMC102582.
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-06-23
编制人:磊子
审稿人:叶凡