孤岛的终结:合成微生物菌群(Synthetic Consortia)的底层交互机制与工程化重构
来源:武汉市灰藻生物科技有限公司 浏览量:93 发布时间:2026-05-31 17:24:59
一、引言:科赫法则的百年禁锢与破局
自 19 世纪末罗伯特·科赫(Robert Koch)确立了微生物的“纯培养”法则以来,分离并利用单一纯种微生物进行发酵,一直是现代工业生物技术的绝对金标准。
通过将目标代谢途径强行塞入大肠杆菌或酿酒酵母等单一底盘细胞中,人类成功量产了胰岛素、青蒿素前体等无数珍贵化合物。
然而,随着合成生物学向更复杂的天然产物与大宗化学品进军,单一细胞工厂的“代谢瓶颈”日益凸显。
当一个细胞被迫同时承担底物降解、辅因子再生以及复杂产物合成时,极易引发严重的代谢负担(Metabolic burden)。
过长的异源途径不仅会耗尽细胞的能量池,中间毒性代谢物的积累更是经常导致宿主细胞的直接死亡。
面对这一工程学极限,科学家们开始将目光重新投向自然界:在真实的土壤、海洋或人体肠道中,没有任何微生物是一座孤岛。
通过借鉴自然生态系统,构建多物种协同作战的合成微生物菌群(Synthetic Microbial Consortia),正在成为下一代生物制造的颠覆性范式。
二、底层逻辑:劳动分工与代谢互养(Cross-feeding)
合成微生物菌群的核心工程学思想在于“劳动分工(Division of Labor)”。
通过将一条冗长、复杂的生物合成途径拆解为多个模块,并分别导入不同的微生物底盘中,可以有效分散单一细胞的代谢压力。
为了确保这些不同分工的菌株能够在同一个发酵罐中稳定共存,而不会因为抢夺营养导致一方被彻底淘汰。
研究人员引入了生态学中的代谢互养(Metabolic Cross-feeding)机制。
最经典的策略是利用营养缺陷型(Auxotrophy)菌株互补:例如,将“菌株 A”设计为无法合成氨基酸 X 但能过量分泌氨基酸 Y;而“菌株 B”恰好相反,无法合成 Y 却能分泌 X。
在强制的互利共生(Obligate mutualism)压力下,A 与 B 必须依赖彼此分泌的代谢物才能生存,从而形成了一个自我锁定的稳定联盟。
这种底层的代谢绑定,是维持人工菌群种群比例动态平衡的最坚实基础。
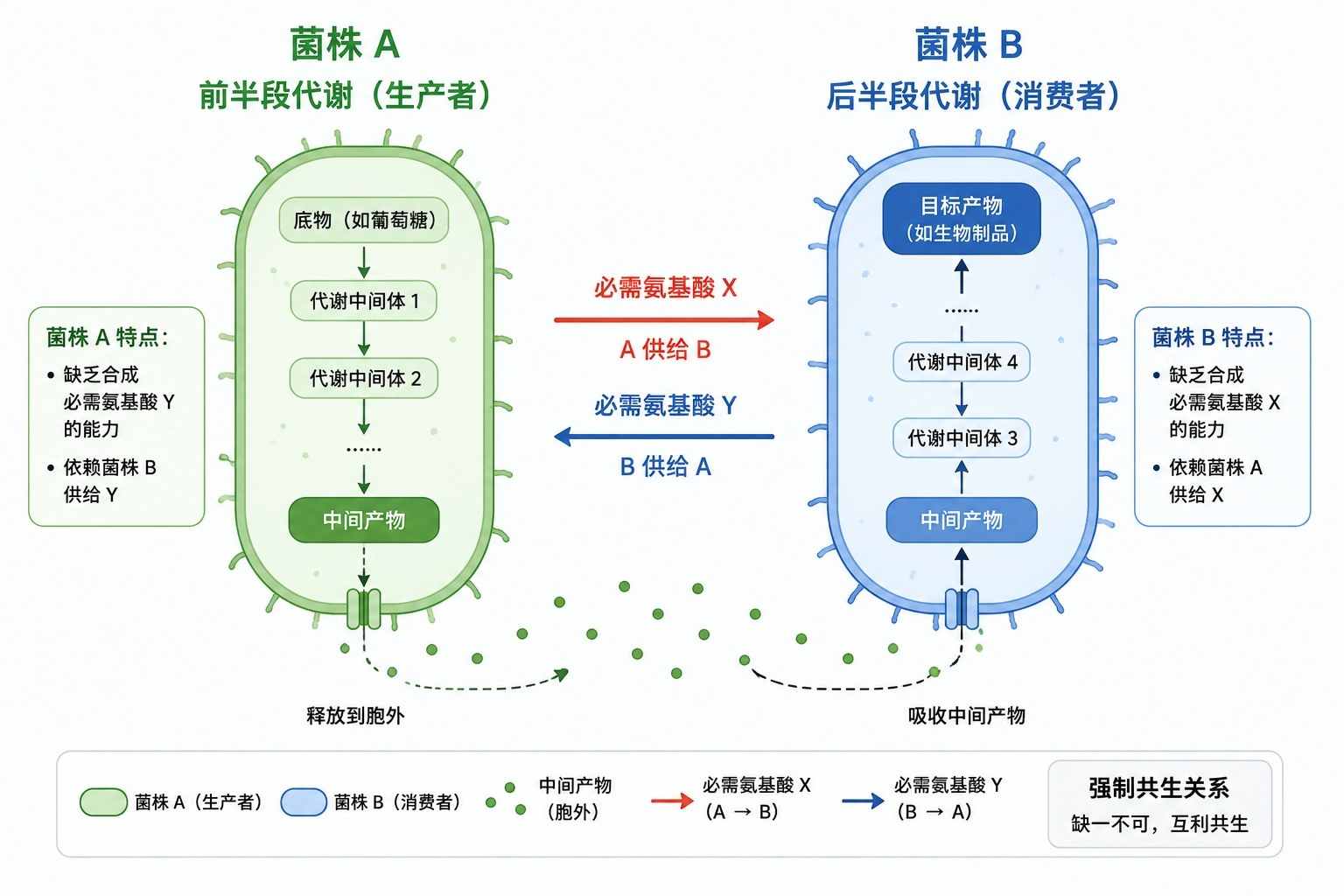
图1. 合成微生物菌群中的空间模块化组装与强制交叉互养机制
三、群落的维稳控制:从群体通讯到动态遗传环路
在实际的工业发酵中,即使建立了代谢互养,不同菌株的生长速率差异依然可能导致群落结构的最终崩溃。
为了实现更高阶的稳定控制,合成生物学家们开始将天然的群体感应(Quorum Sensing, QS)系统改造成人工的“通讯网线”。
通过引入革兰氏阴性菌的 AHL 信号系统或阳性菌的 AIP 信号系统,研究人员可以构建跨物种的遗传控制环路。
例如,当检测到“菌株 A”的生长密度过高(AHL 浓度飙升)时,该信号会直接触发“菌株 B”表达某种生存因子,同时启动“菌株 A”自身的生长抑制基因(如致死蛋白或质粒消除环路)。
这种类似于“负反馈振荡器”的精密设计,使得人工菌群能够根据实时的细胞密度自主调节群体比例,实现了从“被动共生”到“主动微调”的跨越。
四、从底向上:合成菌群的 DBTL 设计周期
构建一个高效的合成微生物菌群,远非将两种菌株简单地混合在一起。
现代微生态工程严格遵循着 Design-Build-Test-Learn(设计-构建-测试-学习) 的迭代周期。
在设计(Design)阶段,研究者严重依赖于全基因组尺度代谢模型(GEMs)与群体生态学动力学方程(如 Lotka-Volterra 模型)。
通过计算机模拟,预测不同微生物在特定碳源下的生长轨迹与代谢通量分配;
在构建(Build)阶段,利用 CRISPR-Cas 等高通量编辑工具敲除冗余通路,精准植入异源模块;
在测试(Test)环节,通过微流控芯片技术结合高通量测序(16S 或宏转录组),实时追踪微反应器内不同菌株的相对丰度波动;
最终将海量数据反馈至学习(Learn)模块,利用机器学习算法进一步优化互养接头的匹配度与初始接种比例。
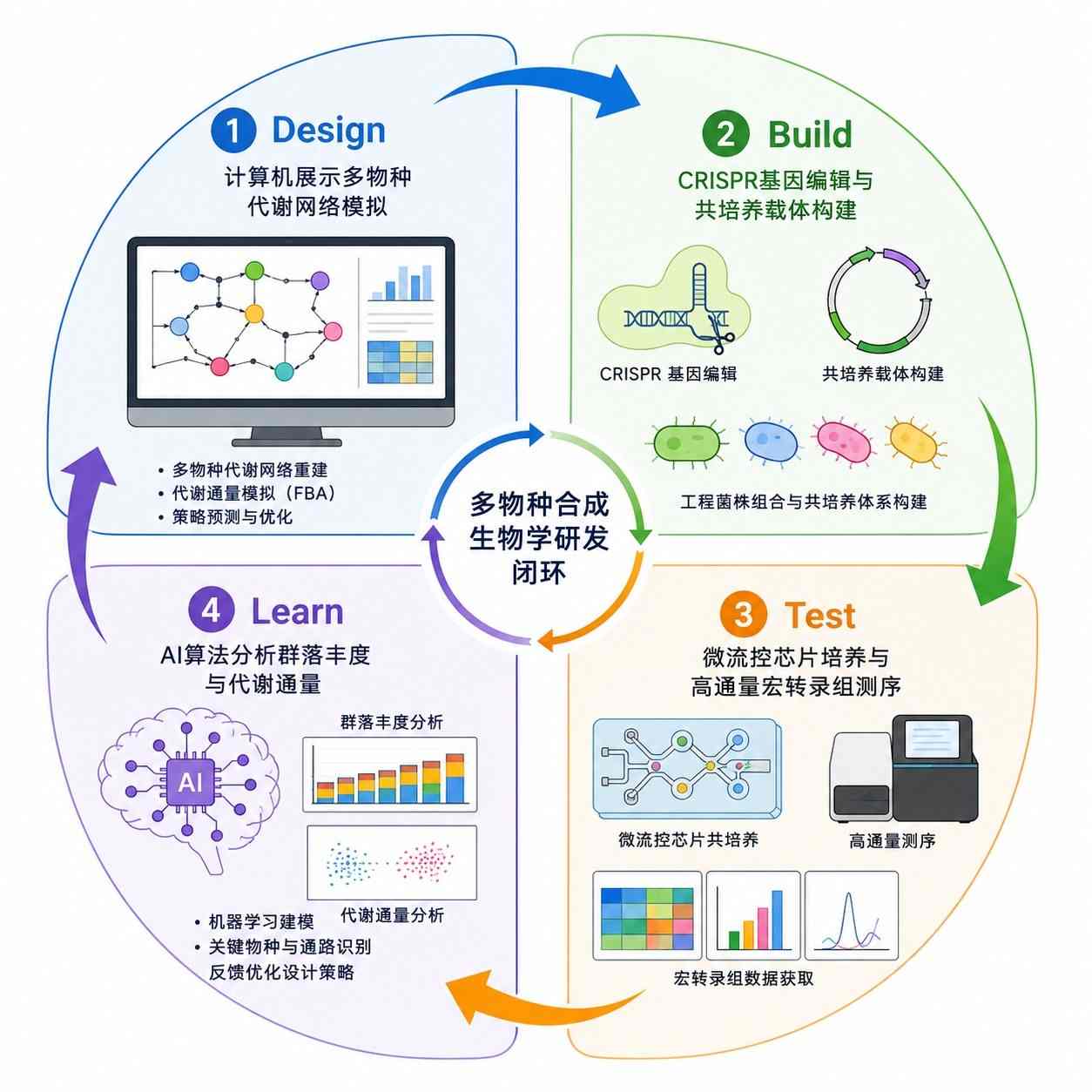
图2. 基于计算生物学与多组学反馈的合成菌群工程化迭代流程
五、前沿应用:从整合生物加工(CBP)到环境修复
人工微生物菌群的成功构建,正在彻底重塑传统工业生物技术的应用边界,并在多个曾被视为禁区的领域展现出极高的商业潜力。
在木质纤维素的生物转化中,传统的工艺需要先用真菌分泌的纤维素酶对秸秆进行昂贵的体外糖化,再由酵母进行发酵。
而通过构建“纤维素降解菌(如解纤维梭菌) + 产物合成菌(如酿酒酵母或大肠杆菌)”的人工菌群,
可以直接实现整合生物加工(CBP),在同一个反应器内同步完成大分子的水解与高价值化学品(如生物丁醇、异丁烯)的合成,极大地降低了生产成本。
在环境生物修复领域,面对土壤和地下水中极其复杂的混合有机污染物(如多环芳烃与重金属的复合污染),
单一微生物往往力不从心。通过设计包含“初级氧化菌”、“脱卤厌氧菌”与“重金属富集菌”的多重微生态群落,
并在其中植入质粒接合转移机制,可以使降解基因在群落内快速横向传播,从而构建出极具环境韧性的超级修复生态墙。
六、结语与展望
从纯培养的孤岛走向复杂协同的微生态网络,合成微生物菌群代表了人类驾驭生命系统能力的又一次飞跃。
在这场微观的社会重构中,不同物种的底盘细胞不再是互相倾轧的竞争者,而是精密机械中咬合紧密的齿轮。
然而,要实现这些人工菌群在工业级发酵罐中长达数月的稳定运行,我们仍需面对质粒丢失、进化漂变等诸多挑战。
对于专业的生物保藏与资源提供机构而言,构建并维持一个遗传背景清晰、表型数据完善的标准底盘细胞库,是支撑这一宏大构想的基础。
未来,随着更多非模式微生物(如极端嗜极菌、专性厌氧菌)被纳入群落设计的拼图,我们必将解锁一个由微生态引擎驱动的绿色制造新纪元。
参考文献
1. Brenner, K., You, L., & Arnold, F. H. (2008). Engineering microbial consortia: a new frontier in synthetic biology. Trends in biotechnology, 26(9), 483-489.
2. Rozen, D. E., et al. (2012). Evolution of interspecific mutualism through experimental co-adaptation of bacteria. Nature, 492(7429), 239-242.
3. D'Souza, G., et al. (2018). Ecology and evolution of metabolic cross-feeding interactions in bacteria. Natural Product Reports, 35(5), 455-488.
标准底盘细胞与共培养参考菌株
HZB221626:枯草芽孢杆菌 (Bacillus subtilis) | 标准表型革兰氏阳性底盘菌
HZB208299:酿酒酵母 (Saccharomyces cerevisiae) EBY100 | 真核共培养优质底盘
敬请关注灰藻生物,共筑健康未来!— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-05-31
编制人:小段
审稿人:叶凡