从基础开始学习原核细胞07:细菌的趋化性与芽孢
来源:武汉市灰藻生物科技有限公司 浏览量:201 发布时间:2026-05-26 13:23:38
一、趋化性 (Chemotaxis)
细菌的运动并非总是随机的。它们会被糖、氨基酸等营养物质吸引,同时也会避开许多有害物质和自身代谢废物。
细菌还能对其他环境信号做出反应,例如温度(趋热性, thermotaxis)、光(趋光性, phototaxis)、氧气(趋氧性, aerotaxis)、渗透压(趋渗性, osmotaxis)和重力。
这种朝向化学引诱物或远离驱避物的运动被称为趋化性 (chemotaxis)。这种行为对细菌显然有利。
可以通过一个实验来演示趋化性:将一根细毛细管装满引诱物,然后插入细菌悬液中。当引诱物从毛细管末端扩散出来时,细菌会聚集并向上游动进入管内。短时间内进入毛细管的细菌数量,反映了引诱物的吸引力强度和趋化速率。
正趋化性和负趋化性也可以在培养皿中进行研究(图1)。如果将细菌置于含有引诱物的半固体琼脂平板中央,细菌会先耗尽局部的营养,然后沿着自己制造的引诱物浓度梯度向外游动,形成一个不断扩大的细菌环。
当把含有驱避物的圆片放在接种了细菌的半固体琼脂平板上时,细菌会游离驱避物,在圆片周围形成一个清晰的无菌区(图2)。
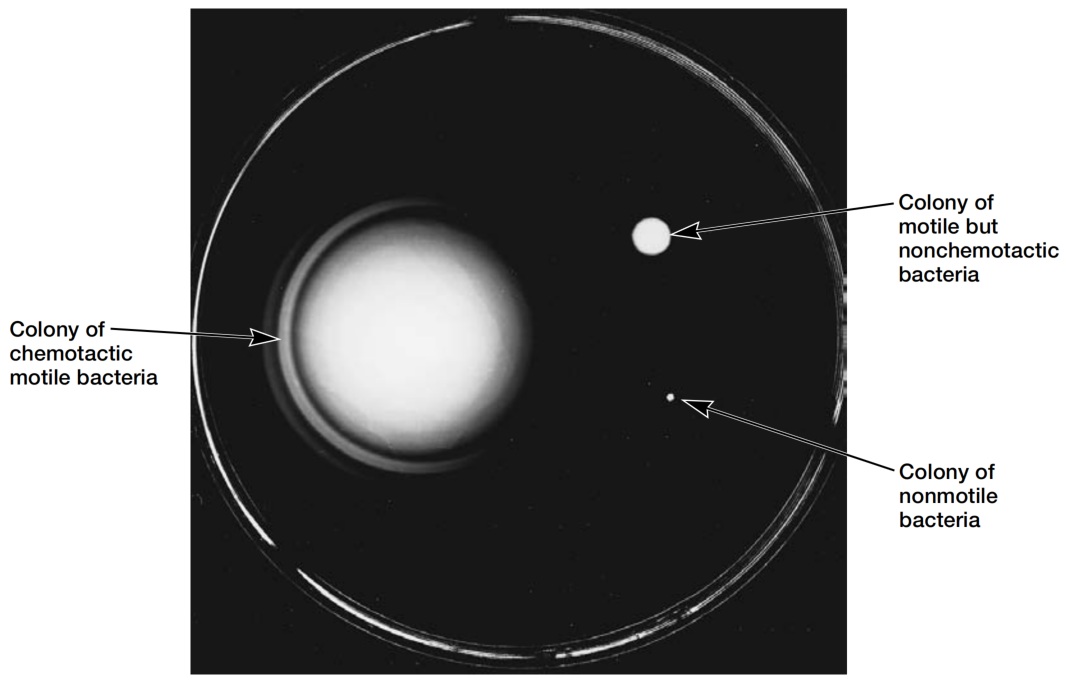
图1 细菌的正趋化性 (Positive Bacterial Chemotaxis)。可在含多种营养物的琼脂平板上观察趋化性。左侧为大肠杆菌 (E. coli) 的正趋化性。外环由消耗丝氨酸 (serine) 的细菌组成。第二环由消耗天冬氨酸 (aspartate, 一种较弱的引诱物) 的大肠杆菌形成。右上角的菌落由能运动但无趋化性的突变体组成。右下角的菌落由不能运动的细菌形成。
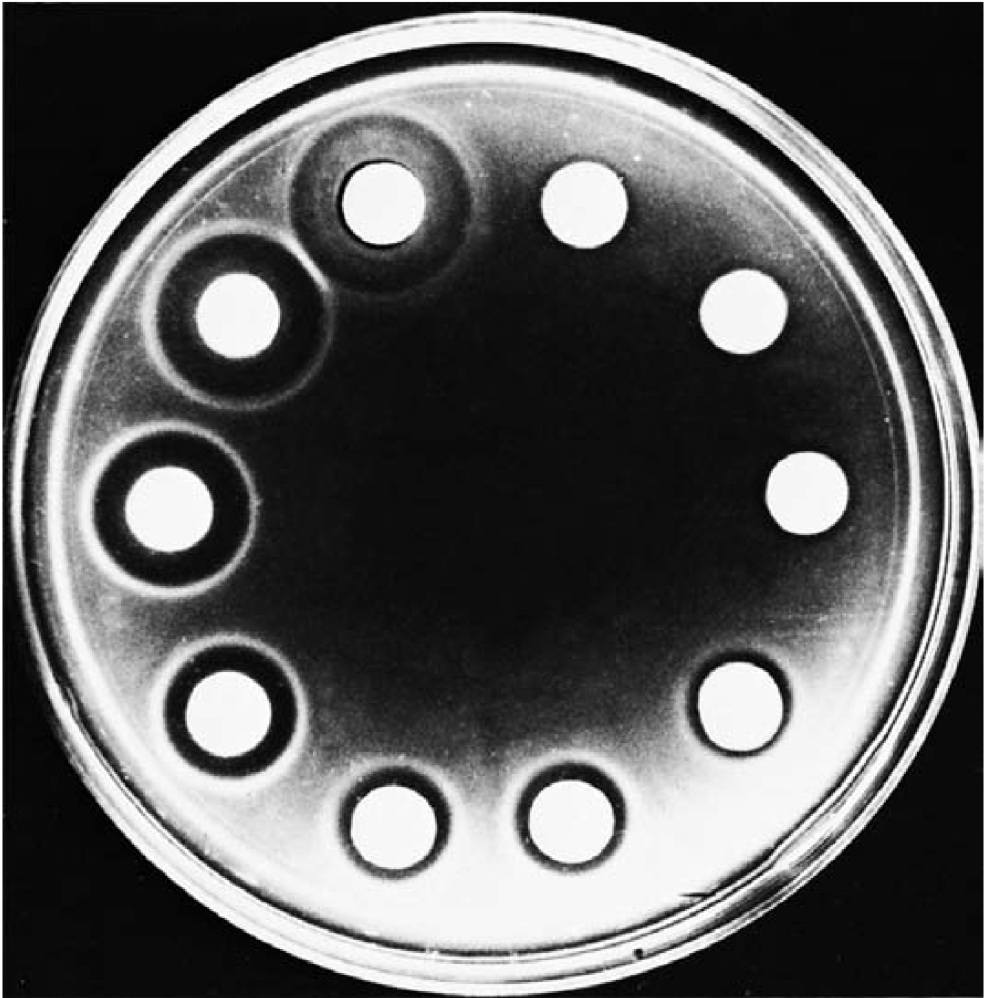
图2 细菌的负趋化性 (Negative Bacterial Chemotaxis)。大肠杆菌 (E. coli) 对驱避物乙酸盐 (acetate) 的负趋化性。亮色圆盘是含有乙酸盐的浓琼脂块,被放置在接种了大肠杆菌的稀琼脂中。乙酸盐浓度从右上角的零增加到左上角的3M。注意随着乙酸盐浓度增加,无菌区的尺寸也随之增大。细菌迁移了30分钟。
细菌能对极低浓度的引诱物(某些糖类约为10-8 M)做出反应,其反应强度随引诱物浓度增加而增大。通常,它们只在较高浓度下才能感知驱避物。
如果引诱物和驱避物同时存在,细菌会比较两种信号,并对有效浓度更高的化学物质做出反应。
引诱物和驱避物由化学感受器 (chemoreceptors) 检测。这些特殊的蛋白质能结合化学物质,并将信号传递给趋化感应系统的其他组分。
目前已发现约20种针对引诱物的化学感受器和10种针对驱避物的化学感受器。这些化学感受器蛋白可能位于周质空间 (periplasmic space) 或细胞质膜 (plasma membrane) 中。有些感受器还参与糖类进入细胞的初始转运阶段。
利用追踪显微镜 (tracking microscope) 可以研究细菌的趋化行为。这种显微镜带有移动载物台,能自动跟踪单个细菌。
在没有化学浓度梯度的情况下,大肠杆菌 (E. coli) 等细菌会随机运动。细菌会先直线或略带弯曲地移动几秒钟,这称为一次“奔跑 (run)”。
当细菌奔跑时,其鞭毛会组织成一个协调的、螺旋状的束。随后,鞭毛束会散开,细菌停止并“翻滚 (tumble)”。
翻滚使细菌随机改变方向,因此下一次奔跑通常会朝向一个新的方向(图3a)。
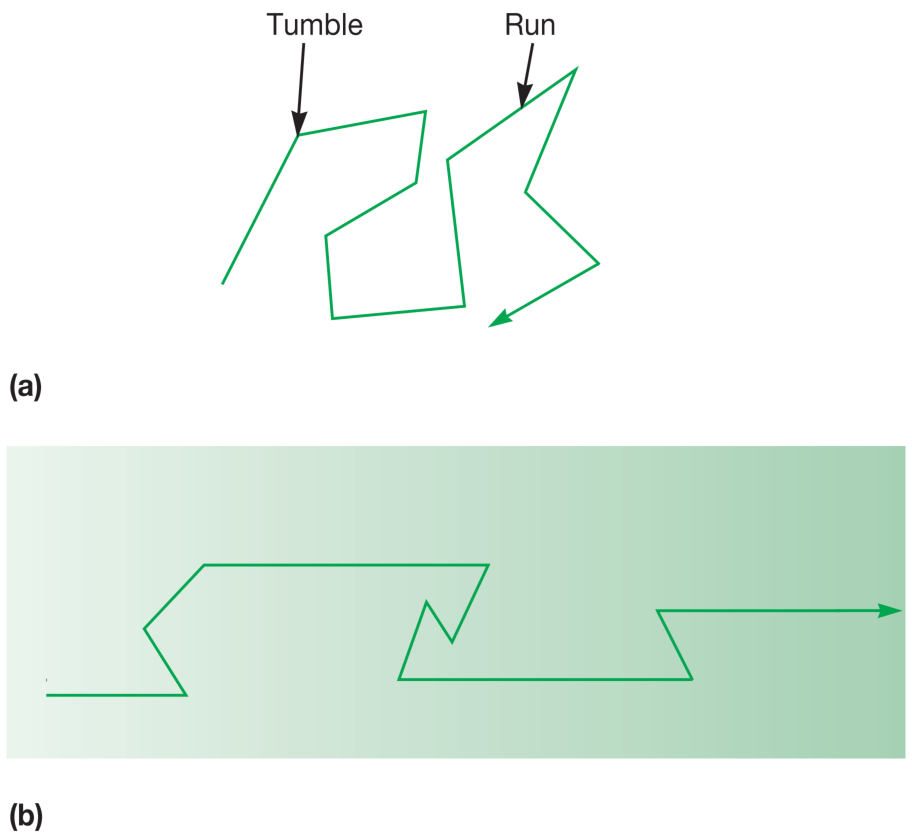
图3 细菌的定向运动 (Directed Movement in Bacteria)。(a) 在无浓度梯度时,细菌的随机运动。翻滚频率相当恒定。(b) 在引诱物梯度中的运动。当细菌向梯度上游(浓度更高处)移动时,翻滚频率降低。因此,朝向引诱物浓度增加方向的奔跑时间更长。
相比之下,当细菌暴露于引诱物中时,如果它正朝着引诱物方向移动,其翻滚频率会降低(或奔跑时间延长)。
尽管翻滚仍可能使细菌偏离引诱物方向,但总体而言,细菌会越来越接近引诱物(图3b)。
对于驱避物,则会产生相反的反应。当细菌远离驱避物时,翻滚频率会降低(奔跑时间延长)。
显然,细菌必须具备某种机制来感知自己正在接近引诱物(或正在远离驱避物)。
细菌的行为由化学浓度的时间变化所塑造。它之所以向引诱物移动,是因为感知到引诱物浓度在增加。同样,它之所以远离驱避物,是因为感知到驱避物浓度在降低。
二、细菌芽孢 (The Bacterial Endospore)
一些革兰氏阳性菌 (Gram-positive bacteria) 能形成一种特殊的、具有高度抗性的休眠结构,称为芽孢 (endospore)。
芽孢在几个属的细菌营养细胞 (vegetative bacterial cells) 内发育,包括芽孢杆菌属 (Bacillus) 和梭菌属 (Clostridium)(杆菌),以及芽孢八叠球菌属 (Sporosarcina)(球菌)等。
这些结构对环境胁迫具有非凡的抵抗力,如高温、紫外线辐射、伽马射线、化学消毒剂和干燥。
事实上,一些芽孢的存活时间可达约10万年。由于其强大的抗性,以及几种产芽孢细菌是危险的病原体,芽孢在食品、工业和医学微生物学中具有重要的实际意义。
这是因为对溶液和固体物品进行灭菌至关重要。芽孢常常能在沸水中存活一小时以上,因此许多材料必须使用高压灭菌锅 (autoclaves) 进行灭菌。
芽孢也具有相当的理论研究价值。因为细菌能在短短几小时内以非常有序的方式制造出这些复杂的结构,所以孢子形成过程非常适合用于研究复杂生物结构的构建。
在自然环境中,当水分或营养物质匮乏时,芽孢有助于细菌生存。
可以使用光学显微镜和电子显微镜检查芽孢。由于芽孢对大多数染料不透,用亚甲蓝 (methylene blue) 等简单染色法处理细菌时,芽孢常呈现为无色区域;需要使用特殊的芽孢染色法才能使其清晰可见。
芽孢在母细胞(即孢子囊, sporangium)中的位置在不同物种间常有差异,这在鉴定上具有重要价值。
芽孢可位于中央、靠近一端(近端, subterminal)或明确位于末端(图4)。有时芽孢体积很大,会使孢子囊膨胀。
图4 芽孢位置与大小的示例 (Examples of Endospore Location and Size)。
电子显微照片显示,芽孢结构很复杂(图5)。芽孢外部常有一层薄而精细的覆盖物,称为外孢衣 (exosporium)。
外孢衣下方是孢子衣 (spore coat),由数层蛋白质构成,可能相当厚。它对许多有毒分子不透,是芽孢抵抗化学物质的主要原因。人们还认为孢子衣含有参与萌发的酶。
皮层 (cortex) 位于孢子衣下方,可占芽孢总体积的一半。它由肽聚糖 (peptidoglycan) 构成,但其交联程度低于营养细胞中的肽聚糖。
芽孢细胞壁(或称核心壁, core wall)位于皮层内部,包裹着原生质体或芽孢核心 (spore core)。核心内含有核糖体 (ribosomes) 和拟核 (nucleoid) 等正常细胞结构,但处于代谢失活状态。
芽孢干重的15%由吡啶二羧酸 (dipicolinic acid) 与钙离子 (calcium ions) 形成的复合物构成(图6),该复合物位于核心内。
长期以来,人们认为吡啶二羧酸直接参与了耐热性,但已分离出缺乏吡啶二羧酸的耐热突变体。
钙确实有助于抵抗湿热、氧化剂,有时也抵抗干热。钙-吡啶二羧酸盐 (calcium-dipicolinate) 可能起到稳定芽孢核酸的作用。
此外,芽孢内还含有一种特殊的、小分子的、酸溶性DNA结合蛋白 (SASPs, small, acid-soluble DNA-binding proteins)。它们能饱和芽孢DNA,保护其免受热、辐射、干燥和化学物质的伤害。
原生质体的脱水 (dehydration) 对耐热性似乎非常重要。皮层可能通过渗透作用从原生质体中移除水分,从而保护其免受热和辐射损伤。
孢子衣也能保护芽孢免受酶和过氧化氢 (hydrogen peroxide) 等化学物质的侵害。
最后,芽孢含有一些DNA修复酶。一旦芽孢萌发,细胞恢复活性,就会对DNA进行修复。
总之,芽孢的耐热性可能是多种因素共同作用的结果:钙-吡啶二羧酸盐和酸溶性蛋白对DNA的稳定作用、原生质体脱水、孢子衣、DNA修复能力,以及适应高温生长的细菌其细胞蛋白本身具有更高的稳定性等。
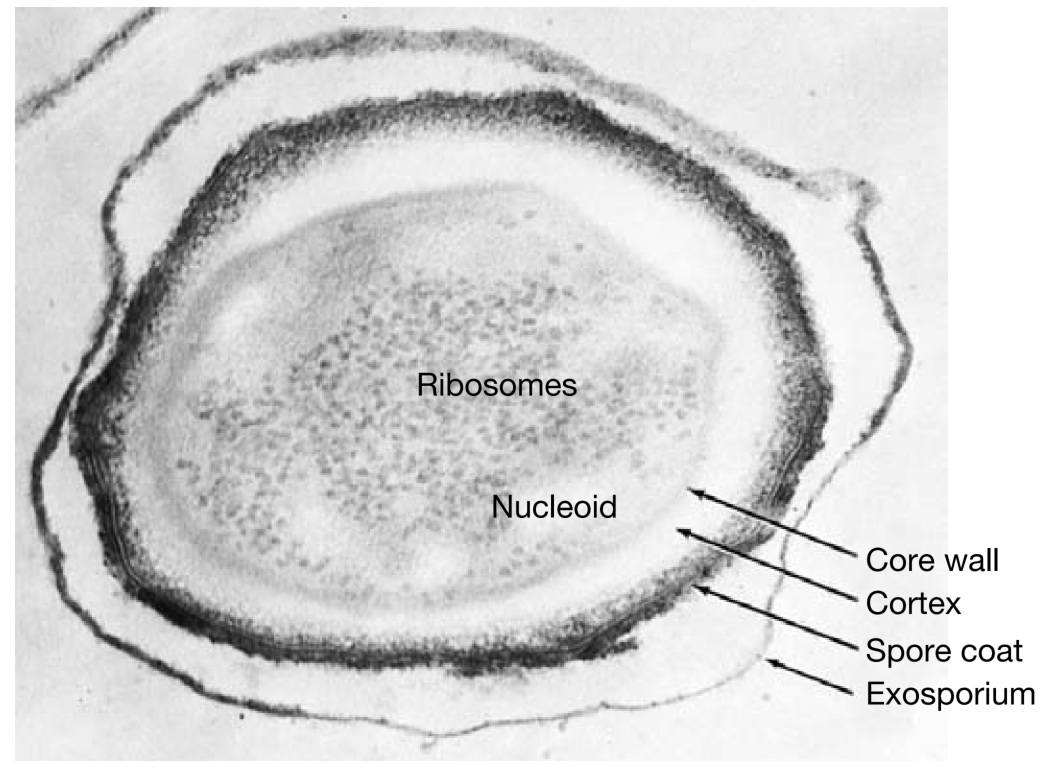
图5 芽孢结构 (Endospore Structure)。炭疽芽孢杆菌 (Bacillus anthracis) 芽孢(×151,000)。
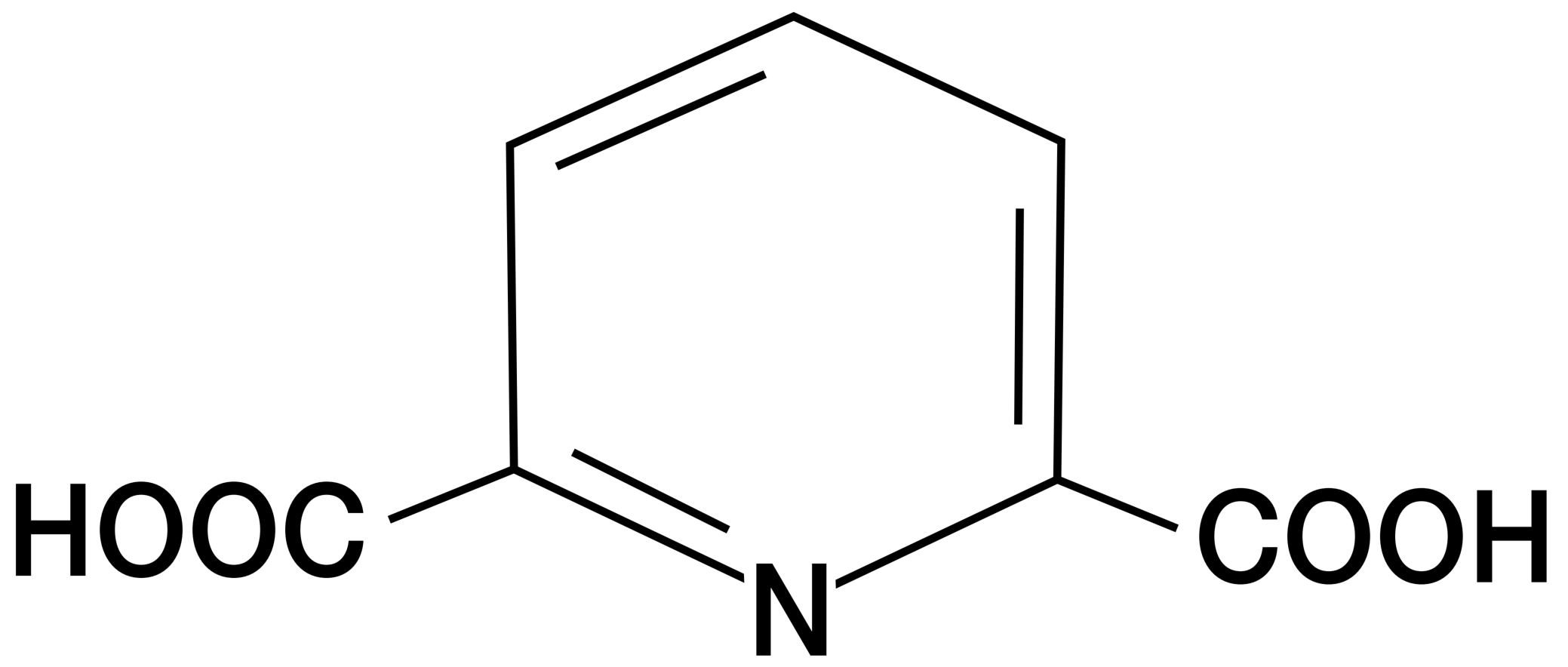
图6 吡啶二羧酸 (Dipicolinic Acid)。
芽孢形成 (Endospore formation),也称为孢子生成 (sporogenesis) 或孢子形成 (sporulation),通常在因缺乏营养而停止生长时开始。
这是一个复杂的过程,可分为七个阶段(图7)。首先形成一条轴向的核物质丝(阶段I),接着细胞膜向内折叠,包裹部分DNA,形成前孢子隔膜(阶段II)。
膜继续生长,将未成熟的芽孢包裹进第二层膜内(阶段III)。随后,在两层膜之间的空间沉积皮层,并积累钙和吡啶二羧酸(阶段IV)。
接着在皮层周围形成蛋白质外壳(阶段V),芽孢进入成熟阶段(阶段VI)。最后,裂解酶破坏孢子囊,释放出芽孢(阶段VII)。
巨大芽孢杆菌 (Bacillus megaterium) 的孢子形成过程大约需要10小时。
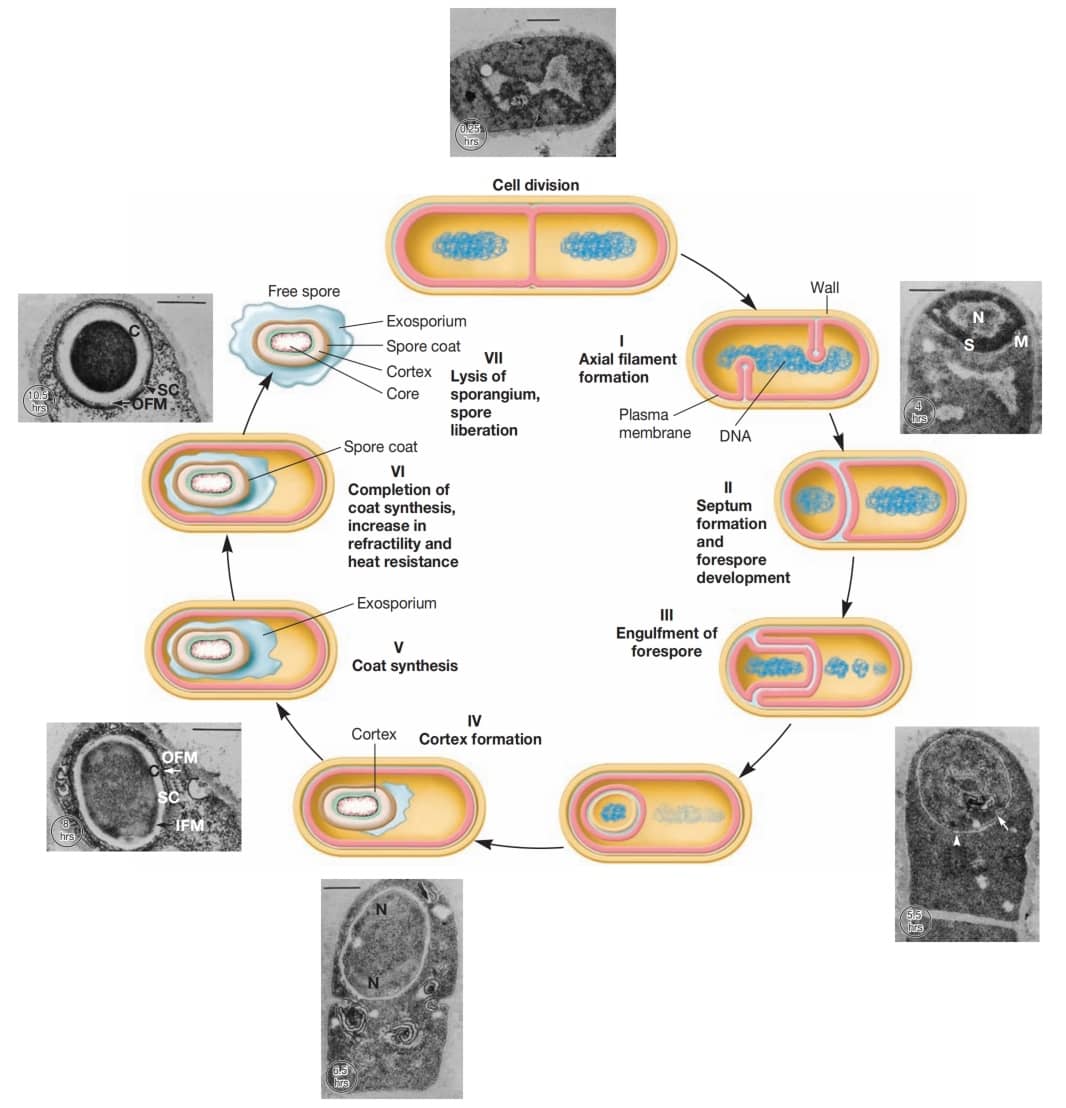
图7 芽孢形成:巨大芽孢杆菌 (Bacillus megaterium) 的生命周期 (Endospore Formation: Life Cycle of Bacillus megaterium)。各阶段用罗马数字表示。照片中的圈内数字指从对数生长期结束起算的小时数:0.25小时—典型的营养细胞;4小时—II期细胞,隔膜形成;5.5小时—III期细胞,吞噬作用;6.5小时—IV期细胞,皮层形成;8小时—V期细胞,外壳形成;10.5小时—VI期细胞,孢子囊内的成熟芽孢。所用缩写:C,皮层;IFM和OFM,前孢子的内膜和外膜;M,间体;N,拟核;S,隔膜;SC,孢子衣。标尺 = 0.5 μm。
休眠芽孢转变为活跃的营养细胞的过程,几乎与孢子生成一样复杂。该过程分为三个阶段:(1) 活化 (activation),(2) 萌发 (germination),和 (3) 生长 (outgrowth)(图8)。
通常,即使在营养丰富的培养基中,芽孢若未经活化也无法成功萌发。活化是一个为萌发做准备的过程,通常由加热等处理引发。
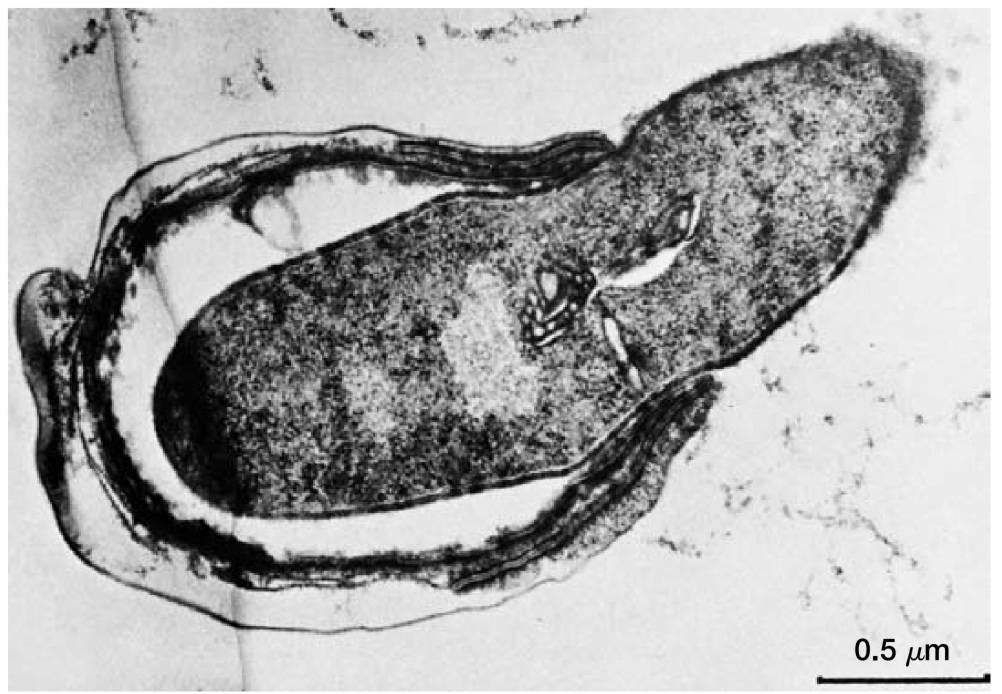
图8 芽孢萌发 (Endospore Germination)。果胶梭菌 (Clostridium pectinovorum) 在萌发过程中从芽孢中伸出。
活化之后是萌发,即打破芽孢的休眠状态。此过程的特点包括:芽孢膨胀、孢子衣破裂或被吸收、对热和其他胁迫的抵抗力丧失、折光性 (refractility) 丧失、芽孢组分释放以及代谢活性增加。
许多正常的代谢物或营养物质(如氨基酸和糖)在活化后都能触发萌发。
萌发之后是第三个阶段,即生长。芽孢原生质体合成新的组分,从残留的孢子衣中伸出,并再次发育成一个活跃的细菌。
参考文献
《Microbiology (Seventh Edition)》 | 微生物学,第七版
相关产品
HZB669242:巴氏孢子八菌DSM 33/Sporosarcina pasteurii(ATCC 11859)
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-05-26
编制人:叶凡
审稿人:小藻