盐碱土壤微生物功能特性与应用综述
来源:武汉市灰藻生物科技有限公司 浏览量:132 发布时间:2026-05-22 14:09:47
一、 引言
盐碱地微生物是生态修复的关键资源。本文综述了其适应机制、群落构建规律及农业应用潜力。微生物通过基因调控、渗透调节等策略应对盐碱胁迫,并受环境过滤与随机过程共同组装。植物促生根际细菌(PGPR)通过促生、抗逆及养分活化等机制提升作物耐盐性。相比传统改良措施,微生物修复更具环境友好性与可持续性。未来需攻克田间定殖难题,开发定制化合成群落,以推动盐碱地绿色高效利用。
二、 盐碱土壤微生物的适应机制
盐碱环境对生物体是巨大的生理胁迫。为了在高盐、高碱的环境中生存,微生物演化出了从基因到代谢层面的精密防御体系。
1. 微生物物种的多样性与分类
盐碱土中的微生物群落复杂多样,主要包括耐盐(能在低盐/无盐下生长,也能耐受高盐)和嗜盐(需要盐分才能生长)两大类。
• 主要类群: 常见的耐盐益生菌主要包括芽孢杆菌属(Bacillus)、假单胞菌属(Pseudomonas)和葡萄球菌属(Staphylococcus)。
• 分类梯度: 根据Kushner和Kamekura 的分类,嗜盐菌分为四类:非嗜盐菌(<0.2 M NaCl)、轻度嗜盐菌(0.2–0.5 M)、中度嗜盐菌(0.5–2.5 M,盐生环境大多数属于此类)和极端嗜盐菌(>2.5 M)。
• 特殊案例: 如黑酵母 Hortaea werneckii 能在无NaCl到饱和NaCl条件下生长;嗜碱微生物通常在pH 10左右生长最佳。
2. 基因表达的精准调控
不同类型的微生物采用了不同的策略来应对高盐胁迫:
• 细菌(快速精准): 依赖双组分系统(TCS)(如EnvZ/OmpR)感知渗透压变化,快速调节基因表达。同时,通过σ因子和翻译后修饰(如赖氨酸乙酰化)来精细调控渗透保护物质的合成。
• 古菌(全局重组): 采用“生存第一”策略,通过全局转录重编程,优先将能量用于维持渗透平衡,而暂时抑制DNA复制和生长相关的代谢。
• 真菌(网络化响应): 依赖HOG(高渗甘油)通路作为核心信号枢纽,诱导甘油合成和细胞壁重塑。此外,小RNA(miRNA/siRNA)在转录后水平提供快速的基因沉默调控。
3. 生理与代谢的适应性重塑
• 渗透调节与能量策略: 微生物会积累相容性溶质(如海藻糖、甜菜碱)。从生物能量学角度看,它们优先从环境摄取这些物质(如甜菜碱),而非从头合成,以降低代谢成本。例如,铜绿假单胞菌在高盐下会积累不饱和脂类。
• 离子稳态: 通过Na⁺/H⁺反向转运蛋白排出钠离子,并利用Kdp(高亲和力ATP酶)、Trk和Tup系统精细调节钾离子的摄入,维持高K⁺/Na⁺比值。
• 抗氧化防御: 通过上调超氧化物歧化酶(SOD)、过氧化氢酶(CAT)等酶的活性,清除活性氧(ROS)。
• 蛋白质与细胞结构稳定性: 嗜盐微生物的胞内蛋白富含酸性氨基酸,形成亲水层;同时分泌胞外聚合物(EPSs),形成物理屏障并促进土壤团聚体形成。
• 极端环境适应性:嗜盐微生物在基因上已适应了极端的太阳辐射、离子强度和干燥环境,因此是药物研发的潜在理想候选者(图1)。
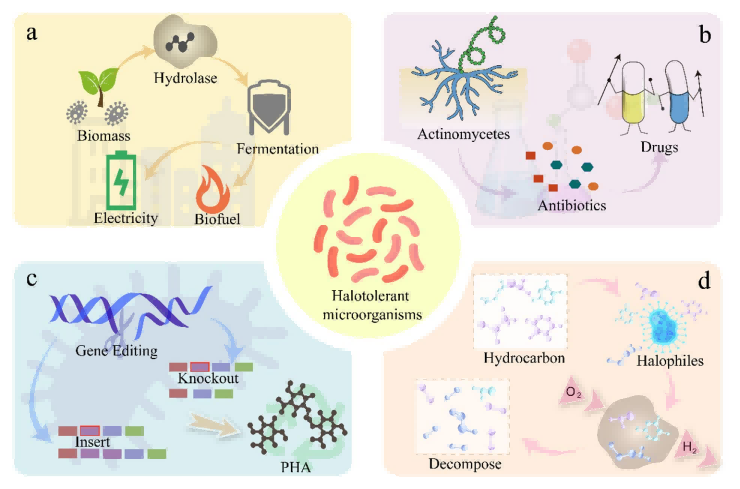
图1. 耐盐微生物的多功能应用。(a) 耐盐微生物利用生物质生产生物燃料和电力。(b) 作为新型抗生素来源的耐盐放线菌。(c) 通过基因编辑技术增强嗜盐微生物的生物产量。(d) 嗜盐微生物在有氧条件下降解碳氢化合物。
三、 盐碱微生物群落的构建机制
微生物群落并非随机聚集,而是受环境过滤和生态过程共同塑造的。中性群落模型解释了这一动态过程。
1. 确定性过程(环境过滤)
这是主导力量,环境因子像筛子一样筛选出适应的物种:
• 盐度的双重机制: 盐度不仅通过渗透胁迫和离子毒性直接筛选群落,还通过改变土壤容重、碳氮含量间接影响。它促使微生物形成更紧密、更复杂的生态网络,这是压力下的稳定性策略。
• pH的直接影响: 极端pH导致蛋白质变性。研究证实,移除植物直接改变土壤pH,会重塑细菌群落结构。
• 根际互作: 根际细菌群落与土体微生物显著不同。耐盐植物的根系分泌物(碳水化合物、氨基酸)能富集特定的促生菌。
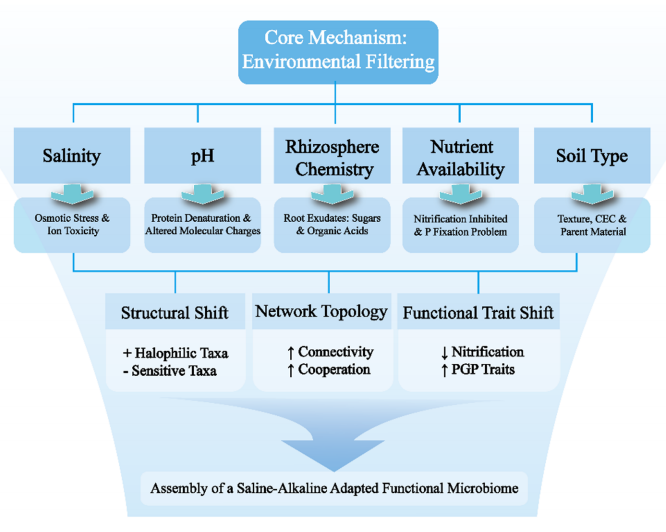
图2.确定性环境过滤塑造了盐碱土壤中的功能性微生物组
2. 随机性过程与中性模型
当环境压力不足以完全抵消随机效应时,中性过程(与功能性状无关)起重要作用:
○ 扩散限制: 即使是微生物,也受地理距离和物理屏障(如山脉、海洋)限制。例如,棉花田地下排水系统中,真菌群落受扩散限制主导。
○ 生态漂变: 在种群数量锐减时(如干旱),随机的遗传漂变会导致群落组成的不可预测性。
○ 平衡框架:
• 胁迫梯度: 在极端高盐(强胁迫)下,确定性过程主导;当盐度降低,随机性过程(如扩散限制)作用增强。
• 尺度依赖: 随机性在深层土壤中影响更大。
• 分类群差异: 真菌群落的组装比细菌更具随机性;稀有类群受确定性选择影响,而丰富类群受随机扩散支配。
表1. 土壤盐渍化的区域驱动因素及其相关特征微生物类群
| 地区 | 气候 | 自然因素 | 人为因素 | 代表性微生物类群 | 关键驱动因素 |
|---|---|---|---|---|---|
| 沙特阿拉伯沙漠地区 | 干旱 | 海水入侵;自然排水不畅 | 水资源管理不善;排水不足 | 极端嗜盐古菌(如 Halobacterium)、Salinibacter ruber、Halorubrum | 极端盐度、高温、强辐射 |
| 中国东部沿海地区 | 季风湿润 | 海岸侵蚀;潮汐盐分入侵;浅层咸地下水 | 土地开垦;工业和生活污水排放 | Salinibacter、Vibrio、海洋变形菌纲(Marine Gammaproteobacteria)、硫酸盐还原菌(如 Desulfovibrio) | NaCl 盐度、潮汐影响、有机污染 |
| 中国西北内陆地区 | 极端干旱 | 咸母质;降雨少;盐分积累;排水不畅 | 不科学灌溉;过度施肥;排水不畅 | Halomonas、Bacillus、耐盐放线菌(如 Streptomyces)、耐干真菌(如 Aspergillus) | 盐度、极端干旱、低有机质 |
| 纳米比亚 | 半干旱 | 沙质土壤易积盐;高蒸发;降雨少 | 农业灌溉和开发改变排水 | 嗜盐/耐盐放线菌、Rhodovibrio、Halothiobacillus | 沙质结构、高蒸发率 |
| 尼罗河三角洲,埃及 | 干旱和半干旱 | 质地细腻、排水不良的土壤;缺乏尼罗河泛滥 | 过度抽水导致海水入侵;筑坝 | Halomonas、Salgentibacter、耐盐 Alphaproteobacteria | 盐度、土壤质地、农业活动 |
| 印度西北部和华东沿海 | 干旱 | 浅层咸地下水毛细上升 | 使用劣质水;化肥残留 | Planococcus、Oceanobacillus、耐盐根瘤菌 | 高蒸发、地下水盐度 |
| 中国东北 | 半湿润和半干旱 | 碳酸钠积累;季节性降雨 | 过度放牧;灌溉和土地管理不善 | 嗜碱古菌(如 Natronobacterium)、Bacillus pseudofirmus、耐碱假单胞菌(Alkalitolerant Pseudomonas) | 高 pH(碱度)、水位波动 |
| 巴西东北部和南里奥格兰德州 | 半干旱 | 靠近大西洋;海水入侵 | 灌溉不当;过度使用咸水 | Halomonas、Cobetia、海洋来源嗜盐菌 | 海水入侵、灌溉水质 |
| 澳大利亚墨累-达令河谷 | 半干旱 | 河口易受气候/水文变化影响;蒸发 | 上游灌溉和大坝改变流量和冲刷 | 耐盐 β-变形菌(Halotolerant Betaproteobacteria)、嗜盐拟杆菌(Halophilic Bacteroidetes)、硫酸盐还原菌 | 盐度波动、水文变化 |
| 伊拉克中南部 | 干旱 | 不透水底土导致渍水和退化 | 灌溉系统效率低下;过度灌溉 | Halomonas、Chromohalobacter、耐盐硫氧化菌 | 排水不畅、次生盐渍化 |
| 孟加拉国西南沿海 | 季风湿润 | 低海拔;海水入侵沿海含水层和土壤 | 土地规划不当;灌溉/排水系统差 | Vibrio、Salinibacter、好氧不产氧光养菌 (AAnP) | 盐度波动、洪水频率 |
| 哈萨克斯坦中南部 | 干旱 | 降水少;蒸发高;中高度盐渍土 | 伊犁河流域灌溉引起的次生盐渍化 | Halomonas、Salicola、嗜盐古菌、产 EPS 细菌 | 盐度梯度、灌溉制度 |
| 阿根廷南部 | 干旱 | 周期性海水淹没和蒸发 | 改变水文的工程结构(堤道、堤坝) | 嗜盐古菌、Salinibacter、紫色硫细菌 | 周期性淹没、水文改变 |
| 澳大利亚西部 | 干旱 | 地形和地下水流使盐分在剖面中富集 | 土地管理不善;灌溉提高盐度 | Halomonas、Salinibacter、多种嗜盐古菌 | 次生盐渍化、地下水位深度 |
| 美国西部 | 干旱或半干旱 | 海洋来源土壤;咸母岩;高钠含量 | 浅地下水位;灌溉引起的排水负荷 | Halomonas、Bacillus、高渗透压下活跃的放线菌 | 钠毒性、灌溉排水 |
四、 微生物的特殊生物技术潜力(医药与工业)
盐生微生物在医药和工业领域的独特价值,这是常被忽略的重要内容:
○ 抗生素与药物开发: 嗜盐碱性放线菌是发现新型抗生素的宝贵资源库。盐度升高会增加土壤中抗生素耐药基因(ARGs)的丰度,耐盐放线菌是主要传播媒介,但也因此具有独特的代谢途径。
○ 具体活性物质:
• 从嗜碱真菌 Sodiomyces alkalinus 中分离的疏水蛋白 Sa-HFB1,对临床真菌有高效活性(潜在新型抗真菌剂)。
• 嗜盐微生物(如部分 Actinomycetes)对金黄色葡萄球菌、大肠杆菌及多种植物病原真菌(如镰刀菌)具有拮抗作用。
○ 工业酶: 嗜碱真菌能分泌纤维素降解酶,富含抗性降解基因,在生物技术领域有巨大潜力。
五、 益生菌效应:植物促生根际细菌 (PGPR)
植物促生根际细菌(PGPR)是盐碱地修复的关键。它们通过多种机制直接促进植物生长并缓解盐胁迫。
1. 调节植物激素
PGPR能分泌吲哚-3-乙酸 (IAA)、赤霉素等,促进根系发育(增加侧根和根毛),从而扩大植物吸收水分和养分的面积。同时,它们能产生ACC脱氨酶,降低植物应激激素乙烯的水平,减少盐胁迫引起的生长抑制。
2. 维持离子平衡与渗透调节
• 脯氨酸与藻酸: 接种耐盐细菌后,玉米的脯氨酸产量提高了55%至191%,藻酸产量增加,有效抵消盐害。
• 排钠保钾: 菌株 Kocuria rhizophila Y1 接种玉米后,显著降低了Na⁺含量和电解质渗漏。
• 激素调节: 菌株 Bacillus BSE01 接种鹰嘴豆,提高了K⁺:Na⁺比值和脯氨酸含量,同时维持较低的ABA(脱落酸)水平。
3. 增强抗氧化与基因表达
○ 基因层面: 接种PGPR后,植物体内的耐盐基因(如 ZmNHX1, ZmDREB2A)和抗氧化基因( ZmGR1, ZmAPX1)转录水平显著上调。
○ 协同作用:
• PGPR + EPS: Peribacillus castrilensis N3 与胞外多糖 Mauran 联用,协同提升番茄耐盐性。
• PGPR + AMF: PGPR产生的ACC脱氨酶促进了丛枝菌根真菌(AMF)的发育,进一步帮助减少钠积累。
4. 改善土壤养分与微环境
• 养分活化: PGPR能溶解难溶性磷、固定大气氮,提高土壤肥力。
• 酶活性提升: 接种PGPR能显著提高土壤中脲酶、磷酸酶、蔗糖酶等的活性,促进有机质分解和养分循环。
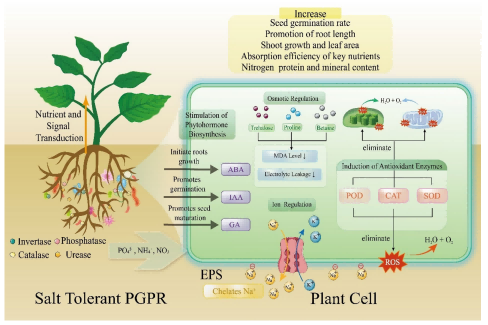
图3.盐胁迫下耐盐植物促生根际细菌(PGPR)促进植物生长的机制。(1) 养分改善与信号转导(2) 植物激素调节(3) 渗透调节(4) 离子稳态(5) 抗氧化防御系统激活
表2.植物促生根际细菌(PGPR)菌株及其缓解不同植物盐胁迫的机制实例
| 作物 | PGPR | 主要机制 | 作物类型 |
|---|---|---|---|
| 拟南芥 (Arabidopsis thaliana L.) | 伯克霍尔德氏菌 (Burkholderia sp.) BK01 | 增加脯氨酸含量,提高 SOD、POD 和 CAT 活性,降低 MDA 含量,有效提高 K⁺/Na⁺ 比率并清除 ROS | 模式植物 |
| 燕麦 (Avena sativa) | PGPR 菌株 IG 3 | 改善幼苗长度、根长、枝条干重、根干重和相对含水量等生理参数,降低蛋白质含量,并正向调节 rbcL 和 WRKY1 的表达谱 | 谷类作物 |
| 甜菜 (Beta vulgaris L.) | SRT15 和 HPJ40 | 减少电解质渗漏和 Na⁺ 吸收,改善叶绿素 a 荧光参数、叶绿素和类胡萝卜素浓度、气孔导度及抗氧化能力 | 糖料作物 |
| 油菜 (Brassica napus L.) | 肠杆菌 (Enterobacter sp.) S16-3 和 假单胞菌 (Pseudomonas sp.) C16-20 | 减少 ROS 诱导的损伤,提高抗氧化酶(如 SOD)的活性,增加生长激素和脯氨酸含量 | 油料作物 |
| 棉花 (Cotton) | 产气肠杆菌 (Enterobacter cloacae) Rs-35 | 促进细胞离子浓度平衡,稳定 IAA 水平的细胞内变化,并刺激系统耐受性的诱导 | 纤维作物 |
| 黄瓜 (Cucumber) | 蜡样芽孢杆菌 (Bacillus cereus) BC56 | 提高幼苗过氧化物酶和谷胱甘肽还原酶活性,清除活性氧,降低脱落酸水平,并诱导光合作用、植物激素、转录调控、代谢物合成与代谢及细胞成分相关基因的表达变化 | 蔬菜作物 |
| 草莓 (Fragaria ananassa) | 枯草芽孢杆菌 (Bacillus subtilis) EY2, 萎缩芽孢杆菌 (Bacillus atrophaeus) EY6, 等 | 促进生长,增加叶绿素含量和养分核生产,减少电解质渗漏 | 水果作物 |
| 大豆 (Glycine max L.) | 日本慢生根瘤菌 (Bradyrhizobium japonicum) USDA 110 与耐盐 恶臭假单胞菌 (Pseudomonas putida) TSAU1 | 促进生长,增加氮和磷含量及可溶性叶蛋白含量,并增加结瘤性状 | 谷类作物 |
| 大豆 (Glycine max L.) | 金龟子链霉菌 (Streptomyces lasalocidi) JCM 3373 | 调节应激反应基因 (GmISP, GmPHD2, GmWRKY54) 和根生长相关基因 (GmPIN1a, GmPIN2a, GmUCCA5, GmYUCCA6) 的表达 | 谷类作物 |
| 陆地棉 (Gossypium hirsutum var. Jin668) | 枯草芽孢杆菌 (Bacillus subtilis) 和 pumilus | 诱导基因的大幅上调和下调、植物-病原体相互作用及植物激素变化 | 纤维作物 |
| 向日葵 (Helianthus annuus L.) | 巴西固氮螺旋体 (Brazilian nitrogen-fixing spirochetes) 和 荧光假单胞菌 (Pseudomonas fluoresens) 菌株 Hysun-33 和 Hysun-39 共接种 | 提高抗氧化剂(如过氧化氢酶、过氧化物酶和抗坏血酸过氧化物酶)的活性以及脯氨酸水平,并降低过氧化氢水平 | 油料作物 |
| 大麦 (Hordeum vulgare L.) | Sicibacter sp. C2 | 诱导磷酸盐溶解,产生吲哚乙酸、铁载体、氰化氢和不同水解酶(蛋白酶、淀粉酶、纤维素酶和脂肪酶),并激活一部分应激反应基因,包括 HVA1, HvDREB1, HvWRKY38, 和 HvP5CS | 谷类作物 |
| 紫花苜蓿 (Medicago sativa L.) | 热带芽孢杆菌 (Bacillus tropicus) YJ33 | 增加生物量产量、株高、抗氧化酶活性和粗蛋白含量,促进整体生长并降低 MDA 含量 | 饲料作物 |
| 薄荷 (Mentha piperita) | 里莫斯链霉菌 (Streptomyces rimosus) C-2012 和 莫诺霉素链霉菌 (Streptomyces monomycini) C-2012 | 促进植物生长,增加总精油和薄荷醇含量,改善叶面营养吸收并增强抗氧化机制 | 药用作物 |
| 烟草 (Nicotiana tabacum L.) | 蜡样芽孢杆菌 (Bacillus cereus) TC01 2, 甲基营养型芽孢杆菌 (Bacillus methylotrophicus) TC02 3, 和 解淀粉芽孢杆菌 (Bacillus amyloliquefaciens) TC03 7 | 增加可溶性蛋白和可溶性糖含量、植酸酶活性、K⁺/Na⁺ 比率,以及 3-吲哚乙酸、玉米素核糖苷和赤霉素的水平 | 其他经济作物 |
| 花生 (Peanut) | 麦芽窄食单胞菌 (Stenotrophomonas maltophilia) BJ01 | 保护光合色素,减少电解质渗漏和脂质过氧化,并增加脯氨酸和 H₂O₂ 含量 | 油料作物 |
| 酸浆 (Physalis heterophylla) | 芽孢杆菌 (Bacillus sp.) 和 荧光假单胞菌 (Pseudomonas fluoresens) | 显著增加产量和块茎中活性成分多糖的积累 | 药用作物 |
| 豌豆 (Pisum sativum) | 铜绿假单胞菌 (Pseudomonas aeruginosa) GKP KS2_7 和 枯草芽孢杆菌 (Bacillus subtilis) MBD 133 | 将 ACC 降解为 α-酮丁酸,表现出 ACCD 活性,并增加叶绿素和抗氧化酶含量 | 蔬菜作物 |
| 甘蔗 (Saccharum officinarum L.) | 芽孢杆菌 (Bacillus sp.) ASN-1 厦门 | 产生生长因子、酶、铁载体、HCN、氨和胞外多糖,并溶解磷酸盐 | 糖料作物 |
| 睡茄 (Withania somnifera) | 铜绿假单胞菌 (Pseudomonas aeruginosa) 菌株 RK3 和 RK4 以及 假单胞菌 (Pseudomonas spp.) RK7 | 产生鼠李糖脂,降低汽油毒性并促进生长和抗氧化活性 | 药用作物 |
| 越橘属 (Vaccinium spp.) | 9株 假单胞菌属 (Pseudomonas spp.) 和 丁基氧化菌属 (ButylOxera spp.) PGPR 菌株 | 增加分枝数、叶片数、叶绿素含量和株高 | 果树作物 |
| 黑吉豆 (Vigna mungo (L.)) | 嗜根考克氏菌 (Kocuria rhizophila)、贝莱斯芽孢杆菌 (Bacillus velezensis) 和 扎根科萨克氏菌 (Kosakonia radicincitans) | 提高光合速率、叶绿素含量、地上部长度和籽粒产量,并分别降低过氧化氢酶和超氧化物歧化酶的酶活性 7.0% 和 1.5% | 谷物作物 |
| 绿豆 (Vigna radiata L.) | 日本慢生型 PGPR TMB7 | 分别使脯氨酸、甜菜碱和总可溶性蛋白含量增加 77%、78% 和 64% | 谷物作物 |
| 葡萄 (Vitis vinifera L.) | 利莫斯链霉菌 (Streptomyces rimosus, Sr) | 改善枝条和根的形态特征,并增加可溶性碳水化合物、可溶性蛋白和脯氨酸含量 | 果树作物 |
| 玉米 (Zea mays L.) | 产碱菌属 (Alcaligenes sp.) E1 和 E2 | 显著改善整体生长特征、抗氧化活性和光合色素含量 | 谷物作物 |
| 枣 (Ziziphus jujuba Mill.) | 解淀粉芽孢杆菌 (Bacillus amyloliquefaciens) CZ-6 | 产生 IAA(生长素)和 ACC 脱氨酶,并分泌纤维素酶、蛋白酶和木聚糖酶,抑制病原真菌的生长 | 果树作物 |
六、 微生物修复与其他改良措施的比较
目前的盐碱地改良策略主要包括物理、化学、水利和生物措施。微生物修复(生物修复的一种)具有独特的优势。
1. 传统改良措施的局限性
表3.盐碱地改良主要措施及其优缺点对比
| 措施类型 | 主要手段 | 优缺点分析 |
|---|---|---|
| 物理改良 | 深耕、客土、覆盖 | 优点: 效果直接,能改善土壤结构。 缺点: 工程量大、成本高,且难以从根本上解决盐分问题。 |
| 化学改良 | 石膏、酸性物质、调理剂 | 优点: 能快速降低pH和置换钠离子。 缺点: 可能引入二次污染,长期使用成本高,对土壤生物有害。 |
| 水利改良 | 灌排、滴灌、咸水利用 | 优点: 利用水分淋洗带走盐分。 缺点: 依赖大量淡水资源,在缺水地区难以实施。 |
2. 微生物修复的独特优势
微生物修复(尤其是利用PGPR和嗜盐微生物)被视为一种环境友好、可持续且低成本的策略:
• 多功能性: 微生物不仅能耐受高盐,还能通过代谢活动(如产酸、产胞外多糖)直接改善土壤理化性质。
• 协同增效: 微生物可以与植物形成共生关系(如联合接种丛枝菌根真菌AMF),比单一的植物修复更高效。
• 生物技术潜力: 嗜盐放线菌和真菌是新型抗生素、抗真菌药物的潜在来源(如从Sodiomyces alkalinus中分离出的抗真菌蛋白)。
七、 从实验室到田间的挑战与展望
尽管PGPR在实验室(温室)条件下表现出色,但将其转化为田间生产力仍面临“死亡之谷”。
1. 主要挑战
• 定殖竞争(Establishment): 盐碱土中原有的土著微生物群落非常强大。外源接种的PGPR菌株往往缺乏趋化性(对根系分泌物的响应)、生物膜形成能力,以及对本地抗生素的耐受性,导致难以在根际成功定殖。
• 制剂生产的难点: 高盐环境本身对细胞有压力。干燥、高温暴露和储存氧化会导致菌体活力丧失。需要添加海藻糖、淀粉、甘油等作为保护剂。
• 施用方式: 种子包衣、沟施或滴灌的效果差异巨大,需根据作物和灌溉习惯优化。
2. 未来方向
✅ 量身定制: 开发基于特定区域土壤和作物的“定制微生物组合群”(SynComs),而非单一菌株。
✅ 多组学应用: 利用多组学方法揭示真实条件下的互作机制。
八、总结
盐碱土壤中的微生物不仅是极端环境的生命奇迹,更是农业可持续发展的关键资源。通过深入理解其适应机制和益生功能,开发高效的微生物肥料,是实现盐渍土生态修复与粮食安全双重目标的必由之路。
参考文献
1.Meinzer M, Ahmad N, Nielsen BL. Halophilic plant-associated bacteria with plant-growth-promoting potential. Microorganisms. 2023;11:2910.
2.Wang P, Liu Q, Fan S, Wang J, Mu S, Zhu C. Combined application of desulfurization gypsum and Biochar for improving saline-alkali soils: a strategy to improve newly reclaimed cropland in coastal mudflats. Land. 2023;12:1717.
3.Strunecký O, Shreedhar S, Kolář L, Maroušková A. Changes in soil water retention following biochar amendment. Energy Sources Part A Recover Util Environ Eff. 2025;47:7145–52.
4.Mosaad ISM, Serag AHI, Sheta MH. Promote sugar beet cultivation in saline soil by applying humic substances in-soil and mineral nitrogen fertilization. J Plant Nutr. 2022;45:2447–64.
5.Naz F, Hamayun M, Rauf M, Arif M, Afzal Khan S, Ud-Din J, et al. Molecular mechanism of Cu metal and drought stress resistance triggered by Porostereum spadiceum AGH786 in Solanum lycopersicum L. Front Plant Sci. 2022;13:1029836.
相关产品
HZB123062:阴沟肠杆菌 | Enterobacter cloacae
HZB368881:蜡状芽孢杆菌 | Bacillus cereus
HZB224023:解淀粉芽孢杆菌 | Bacillus amyloliquefaciens
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-05-22
编制人:思琪
审稿人:叶凡