从基础开始学习原核细胞03:细胞质基质、拟核、质粒
来源:武汉市灰藻生物科技有限公司 浏览量:242 发布时间:2026-04-30 16:05:19
一、细胞质基质(The Cytoplasmic Matrix)
细胞质基质是拟核、核糖体和内含物所悬浮的物质,它主要由水构成(细菌质量约70%是水),不含由脂双层(常称单位膜)包裹的细胞器。质膜及其内部所有物质合称为原生质体,细胞质基质是原生质体的主要组成部分。
1、原核细胞骨架(The Procaryotic Cytoskeleton)
在电子显微镜下可以看到,原核生物的细胞质基质中充满了核糖体。多年来,人们认为原核生物因缺乏细胞骨架,其胞质组织水平低于真核细胞。但近期研究发现,细菌中包含与真核细胞骨架元件(微丝、中间纤维和微管)的同源物,同时在古菌中也发现了一种。这些原核细胞骨架丝在结构和功能上都与真核对应物相似,它们参与细胞分裂、将蛋白质定位到细胞特定位置,并决定细胞形状(图1、表1)。
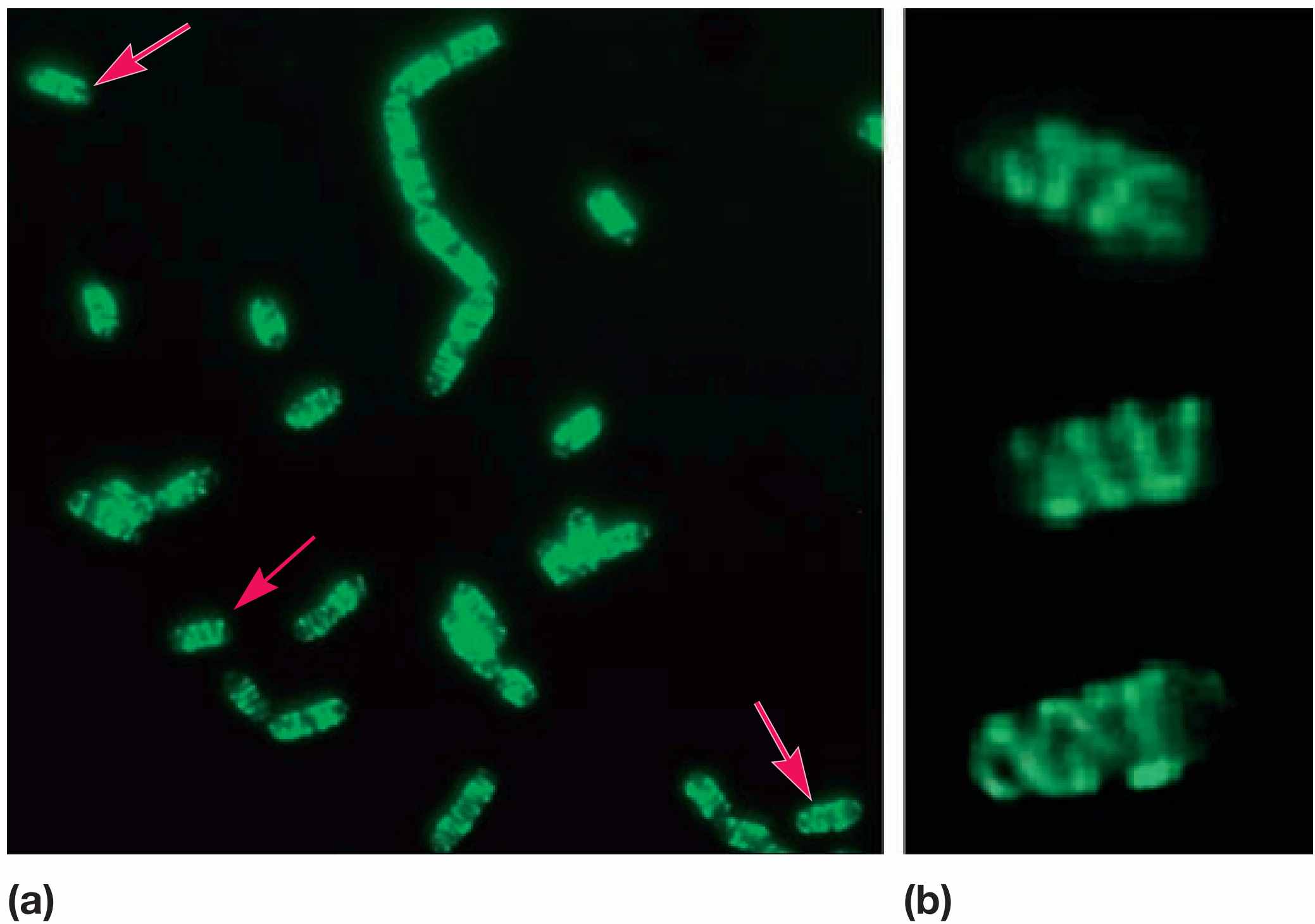
图1 原核细胞骨架。 枯草芽孢杆菌(Bacillus subtilis)中MreB样细胞骨架蛋白(Mbl)的可视化。Mbl蛋白与绿色荧光蛋白融合,通过荧光显微镜观察活细胞。(a) 箭头指向沿细胞全长延伸的螺旋状细胞骨架索。(b)是 (a)中三个细胞的高倍放大图。
表1 原核细胞骨架蛋白(Procaryotic Cytoskeletal Proteins)
| 原核蛋白(真核对应物) | 功能 | 说明 |
| FtsZ(微管蛋白/tubulin) | 细胞分裂 | 在细菌和古菌中广泛存在 |
| MreB(肌动蛋白/actin) | 细胞形状 | 存在于许多杆状细菌中;在枯草芽孢杆菌(Bacillus subtilis)中称为Mbl |
| Crescentin(中间纤维蛋白/intermediate filament proteins) | 细胞形状 | 在新月柄杆菌(Caulobacter crescentus)中发现 |
2、内含物(Inclusion Bodies)
内含物是细胞质基质中的有机或无机颗粒,在光学显微镜下通常清晰可见。它们主要用于储存(如碳化合物、无机物和能量),并通过将分子聚集成颗粒形式来降低渗透压。
一些内含物游离于细胞质中,例如多聚磷酸盐颗粒、蓝细菌素颗粒和部分糖原颗粒。另一些则被一层约2.0至4.0纳米厚的外壳包裹,这层外壳是单层的,可能由蛋白质或蛋白质+磷脂组成的膜状结构构成。被包裹的内含物例子包括聚-β-羟基丁酸(PHB)颗粒、部分糖原和硫颗粒、羧酶体和气泡。许多内含物用于储存,其数量随细胞营养状况而变化。例如,在缺磷的淡水环境中,多聚磷酸盐颗粒会被耗尽。
以下是几种重要内含物的简要描述。
(1)有机内含物,通常有:糖原、聚-β-羟基烷酸酯(如聚-β-羟基丁酸/Poly-β-hydroxybutyrate, PHB)。
糖原是由葡萄糖单元组成的聚合物,其长链通过α(1→4)糖苷键连接,分支链则通过α(1→6)糖苷键连接。
PHB则由β-羟基丁酸分子通过相邻分子的羧基和羟基之间的酯键连接而成。
一个物种通常只含其中一种聚合物,但有些光合细菌两者都有。
PHB会聚集成直径约0.2至0.7微米的独立颗粒,易于被苏丹黑染色用于光镜观察,在电镜下则呈现为空的“孔洞”(见图2-a)。这是因为制备电镜样品的溶剂会溶解这些疏水性内含物。
糖原则以更小的颗粒(直径约20至100纳米)形式更均匀地分散在整个基质中,通常只能在电镜下看到。如果细胞含有大量糖原,用碘液染色会使其呈红棕色。
糖原和PHB内含物是碳储备库,为能量和生物合成提供原料。许多细菌也将碳储存为脂滴。
蓝细菌(一类光合细菌)有两种独特的有机内含物。
蓝细菌素颗粒(见图3-b)由大型多肽组成,其中精氨酸和天冬氨酸含量大致相等。这些颗粒通常足够大,可在光镜下看到,为细菌储存多余的氮。
羧酶体(Carboxysomes)存在于许多蓝细菌和其他固碳细菌中。它们呈多面体,直径约100纳米,内含1,5-二磷酸核酮糖羧化酶(ribulose-1,5-bisphosphate carboxylase),简称Rubisco。Rubisco是固碳的关键酶,负责将大气中的CO2转化为糖。该酶在羧酶体内呈准晶态排列,作为酶的储备库。羧酶体也可能就是CO2固定的场所。
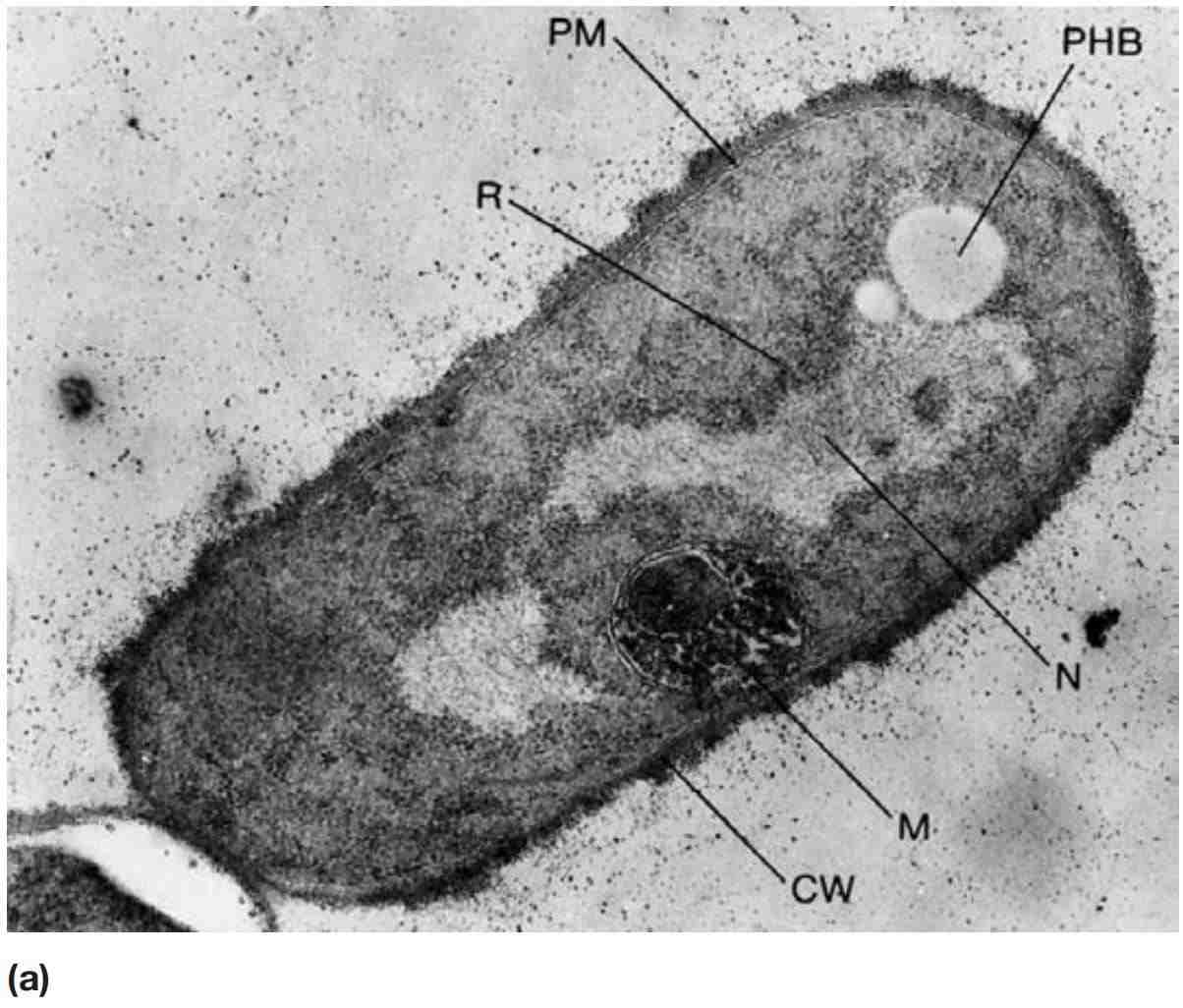
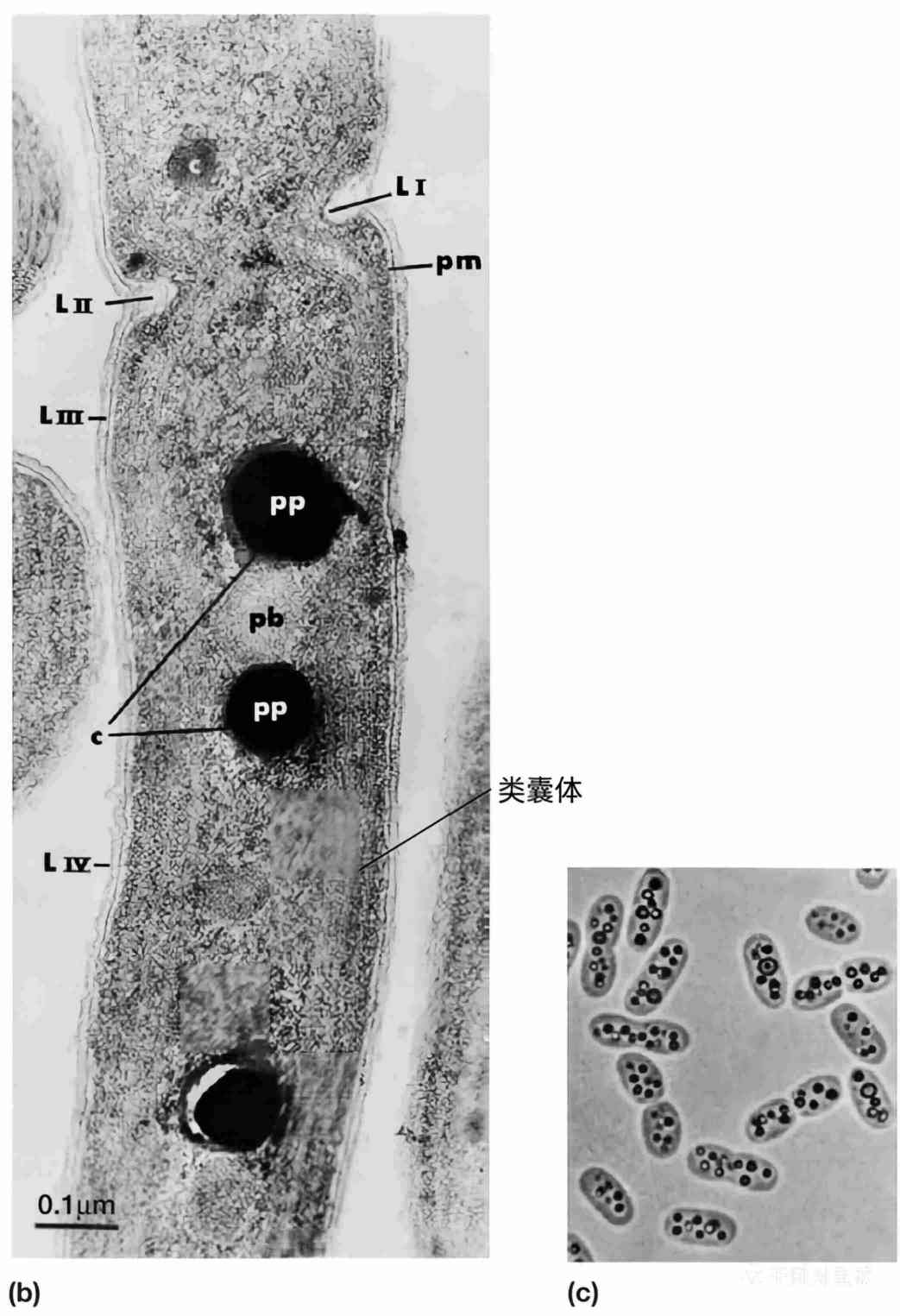
图2 细菌中的内含物。 (a) 巨大芽孢杆菌(Bacillus megaterium)的电镜照片(×30,500)。聚-β-羟基丁酸内含物(PHB);细胞壁(CW);拟核(N);质膜(PM);“间体”(M);核糖体(R)。(b) 集胞藻(Anacystis nidulans)的超微结构。细菌正在分裂,隔膜部分形成(LI和LII)。可见多种结构,包括细胞壁层(LIII和LIV)、多聚磷酸盐颗粒(pp)、多面体(pb)、蓝细菌素物质(c)和质膜(pm)。类囊体(Thylakoids)沿细胞长度分布。(c) 紫色硫细菌——酒色着色菌(Chromatium vinosum)的细胞内硫颗粒,明场显微镜(×2,000)。
一种非常特殊的有机内含物是气泡(gas vacuole),它能为某些水生原核生物提供浮力。
气泡存在于许多光合细菌以及少数其他水生原核生物中,如嗜盐古菌(Halobacterium)和发硫菌(Thiothrix,一种丝状细菌)。气泡由大量微小、中空、圆柱形的结构——气囊(gas vesicles)聚集而成(见图3)。气囊壁完全由一种小型蛋白质构成。这些蛋白质亚基组装成一个坚硬、封闭、中空的圆柱体,不透水但可自由透过大气气体。拥有气泡的原核生物可以调节自身浮力,以悬浮在光线强度、氧气浓度和营养水平合适的深度。它们通过塌陷气囊下沉,通过构建新的气囊上浮。

图3 气囊和气泡。 水华鱼腥藻(Anabaena flos-aquae)的冷冻蚀刻制备样品(×89,000)。雪茄形的气囊聚集成气泡。图中可见气囊的纵向和横截面视图(箭头所示)。
(2)主要的无机内含物:多聚磷酸盐颗粒和硫颗粒。
许多细菌以多聚磷酸盐颗粒(或称异染颗粒/volutin granules)的形式储存磷酸盐(见图2-b)。
多聚磷酸盐是由正磷酸盐通过酯键连接而成的线性聚合物。因此,异染颗粒是磷酸盐的储存库,而磷酸盐是核酸等细胞成分的重要组成部分。在某些细胞中,它们也充当能量储备,多聚磷酸盐可在反应中作为能量来源。这些颗粒有时被称为异染颗粒(metachromatic granules),因为它们具有异染效应:用亚甲蓝或甲苯胺蓝等蓝色染料染色时,会呈现红色或不同深浅的蓝色。
硫颗粒被一些原核生物用于临时储存硫(见图2-c)。
例如,光合细菌可以利用硫化氢作为光合作用的电子供体,并将产生的硫积累在周质空间或特殊的细胞质球状体中。
无机内含物也可用于储存以外的目的。一个很好的例子是磁小体(magnetosome),一些细菌用它来感应地球磁场进行定向。许多这类内含物含有四氧化三铁(磁铁矿/magnetite)形式的铁。
3、核糖体(Ribosomes)
如前所述,细胞质基质通常充满核糖体;它们也可能松散地附着在质膜上。
核糖体是由蛋白质和核糖核酸(RNA)组成的复杂结构。它们是蛋白质合成的场所:细胞质核糖体合成细胞内使用的蛋白质,而质膜上的核糖体则合成运送到细胞外的蛋白质。
新合成的多肽链会在核糖体合成过程中或合成后不久折叠成最终形状。每种蛋白质的形状由其氨基酸序列决定。
一类特殊的蛋白质,称为分子伴侣(molecular chaperones)或简称伴侣蛋白(chaperones),能帮助多肽链正确折叠。
原核核糖体比真核细胞的细胞质或内质网相关核糖体更小。原核核糖体被称为70S核糖体(真核为80S),尺寸约为14-15纳米×20纳米,分子量约270万,由一个50S和一个30S亚基构成。70S中的“S”代表斯维德伯格单位(Svedberg unit),是沉降系数的单位,用于衡量离心机中的沉降速度;颗粒在离心时沉降越快,其斯维德伯格值或沉降系数就越大。沉降系数取决于颗粒的分子量、体积和形状。通常,更重、更紧凑的颗粒具有更大的斯维德伯格数或沉降更快。
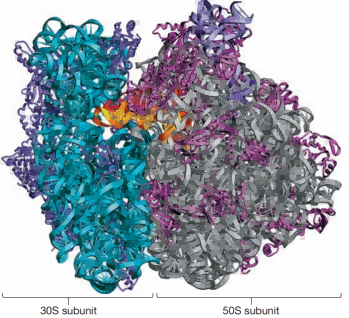
图4 原核细胞核糖体。展示了细菌核糖体的两个亚基。50S亚基包含23S核糖体RNA(灰色)和5S核糖体RNA(浅蓝色),而16S核糖体RNA(深蓝色)则存在于30S亚基中。图中显示了一分子tRNA(金色)位于A位点。为了生成此折线图,需对纯化后的细菌核糖体晶体进行培养,使其暴露于X射线下,并对由此产生的衍射图案进行分析。
二、拟核 (Nucleoid)
原核生物和真核生物最显著的区别在于遗传物质的包装方式。
真核细胞有两条或更多染色体,它们被包裹在由膜界定的细胞核内。而原核生物没有这种膜界定的细胞核,其染色体位于一个不规则的区域,称为拟核 (nucleoid)(也被称为核体、染色质体或核区)(图5)。
通常,原核生物包含一个双链脱氧核糖核酸 (DNA) 的环状分子。但有些细菌具有线性DNA染色体。还有一些细菌,例如霍乱弧菌 (Vibrio cholerae) 和伯氏疏螺旋体 (Borrelia burgdorferi)(分别是霍乱和莱姆病的病原体),拥有多条染色体。
电子显微镜和光学显微镜的研究对于理解拟核的结构和功能非常重要,尤其是在细胞活跃生长和分裂期间。在电子显微照片中,拟核呈纤维状;这些纤维很可能就是DNA。
在活跃生长的细胞中,拟核有延伸到细胞质基质中的突起。这些突起可能包含正在积极转录以产生信使RNA (mRNA) 的DNA。其他研究表明,当遗传物质已经复制但细胞分裂尚未发生时,在单个细胞内可以观察到多个拟核(图5-a)。
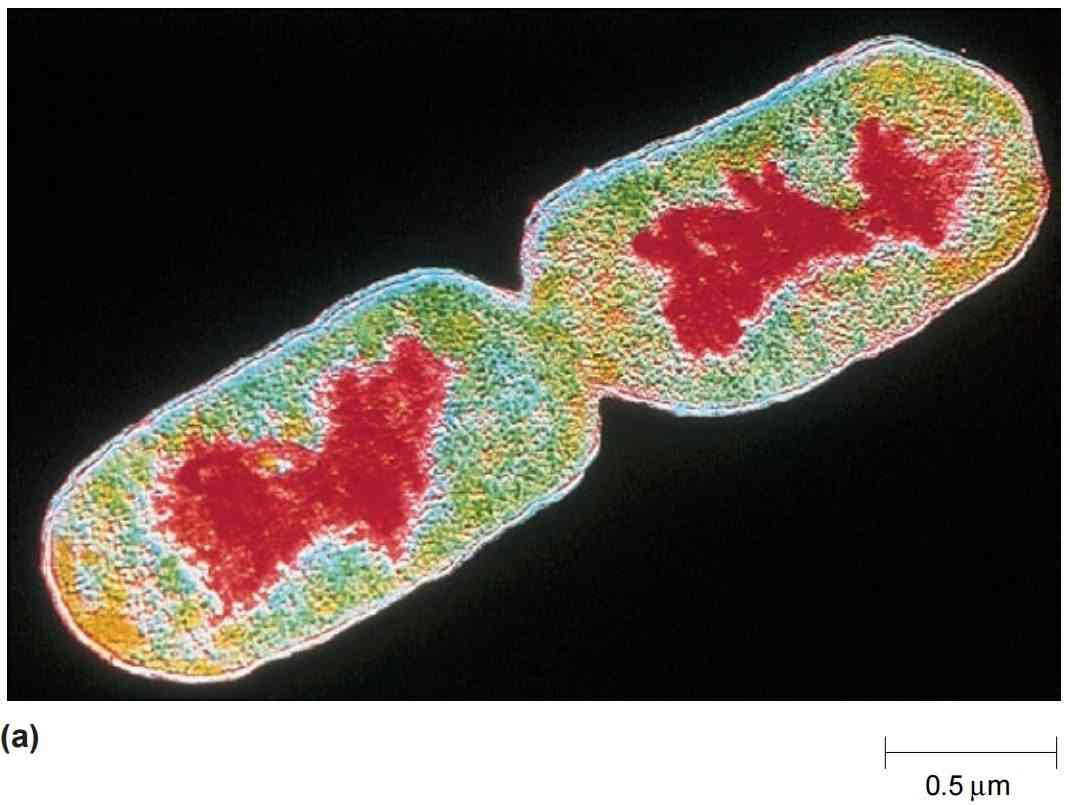
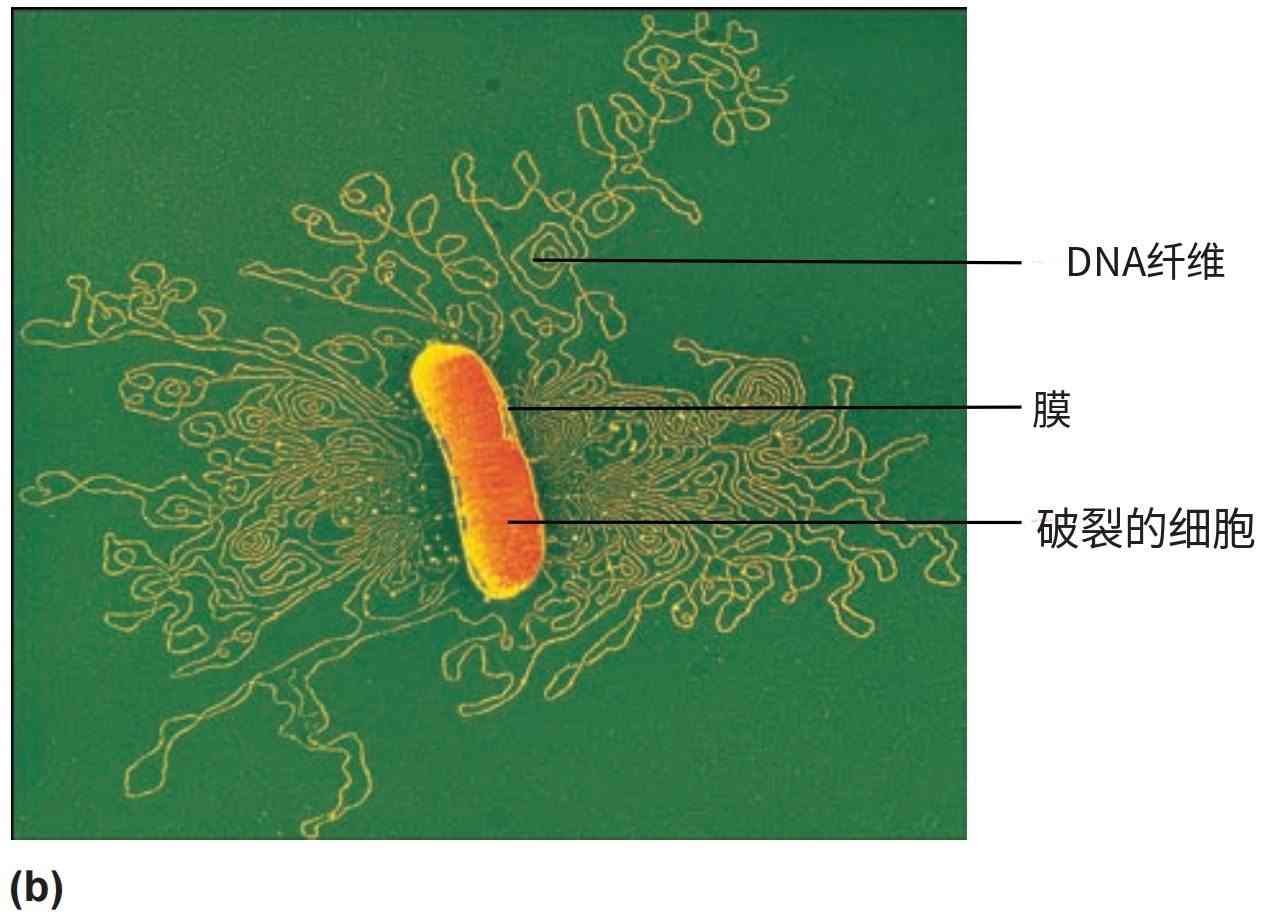
图5 原核生物的拟核和染色体。原核生物的染色体位于细胞质中的拟核区域。(a) 一张经色彩增强的分裂期大肠杆菌 (E. coli) 细胞薄切片透射电子显微照片。红色区域是两个子细胞中存在的拟核。(b) 从温和裂解的大肠杆菌细胞中释放出的染色体。注意DNA在细胞内必须包装得多么紧密。
对纯化拟核的化学分析表明,按重量计算,它们大约由60%的DNA、30%的RNA和10%的蛋白质组成。在大肠杆菌 (Escherichia coli) 中,这个闭合的DNA环长约1400微米,大约是细胞长度的230到700倍(图5-b)。
显然,它必须被非常高效地包装才能适应拟核的空间。DNA被广泛地折叠和缠绕,这可能是在RNA和多种拟核蛋白 (nucleoid proteins) 的帮助下完成的。这些蛋白包括在细菌 (Bacteria) 和古菌 (Archaea) 中都保守的压缩蛋白 (condensing proteins)。与真核生物和一些古菌不同,细菌不使用组蛋白 (histone proteins) 来包装它们的DNA。
浮霉菌门 (Phylum Planctomycetes) 的例外情况
上述情况存在一些例外。在一种特殊的细菌门——浮霉菌门 (Planctomycetes) 的两个属中,存在膜包裹的含DNA区域。
Pirellula(出芽菌属)有一层膜包裹着一个区域,称为pirellulosome,其中包含纤维状的拟核和核糖体样颗粒。Gemmata obscuriglobus(暗球菌属)的核体被两层膜包围。要确定这些膜的功能以及这种现象的普遍程度,还需要更多的研究工作。
三、质粒 (Plasmids)
除了拟核中的遗传物质外,许多原核生物(以及一些酵母和其他真菌)还含有称为质粒 (plasmids) 的染色体外DNA分子。事实上,迄今为止测序的大多数细菌和古菌基因组都包含质粒。
在某些情况下,已在单一物种内鉴定出多种不同的质粒。例如,B. burgdorferi 携带12个线性质粒和9个环状质粒。质粒在其宿主生物的生活中扮演着许多重要角色。它们对微生物学家和分子遗传学家也极为宝贵,可用于构建和转移新的基因组合以及克隆基因。
质粒是小的双链DNA分子,可以独立于染色体存在。已有文献记载了环状和线性质粒,但已知的大多数质粒都是环状的。线性质粒在其末端具有特殊的结构或序列,以防止其降解并允许其复制。质粒的基因数量相对较少,通常少于30个。
它们的遗传信息对宿主并非必需,缺乏质粒的细胞通常也能正常运作。然而,许多质粒携带的基因能在特定环境中为其宿主提供选择性优势。
质粒能够自主复制。单拷贝质粒 (Single-copy plasmids) 在每个宿主细胞中只产生一个拷贝。多拷贝质粒 (Multicopy plasmids) 在每个细胞中的浓度可能达到40个或更多。
一些质粒能够整合到染色体中,因此会随染色体一起复制。这类质粒被称为附加体 (episomes)。质粒在细胞分裂期间能稳定遗传,但它们并不总能平均分配到子细胞中,有时会丢失。质粒的丢失称为消除 (curing)。
消除可以自发发生,也可以通过抑制质粒复制但不影响宿主细胞繁殖的处理来诱导。一些常用的消除处理方法包括吖啶类诱变剂、紫外线和电离辐射、胸腺嘧啶饥饿、抗生素以及高于最适温度的培养。
质粒可以根据其存在方式、传播方式和功能进行分类。
1、接合性质粒 (Conjugative plasmids)
它们拥有编码毛发状结构——菌毛 (pili) 的基因,并能在接合过程中将自身拷贝转移到其他细菌中。
也许被研究得最透彻的接合性质粒是大肠杆菌的F因子(F factor,即致育因子或F质粒),它是第一个被描述的接合因子。F因子包含指导性菌毛形成的基因,这种菌毛能将一个F+细胞(含有F质粒的细胞)连接到一个F-细胞(缺乏F质粒的细胞)上。其他由质粒编码的基因产物有助于DNA从F+细胞转移到F-细胞。F因子还包含几个称为插入序列 (insertion sequences) 的片段,使其能够整合到宿主细胞的染色体中。因此,F因子是一种附加体。
2、抗性因子 (Resistance factors, R factors, R plasmids)
它们赋予细胞抗生素抗性。R因子基因编码的酶能够破坏或修饰抗生素。
一些R质粒只有一个抗性基因,而另一些则多达八个。抗性基因通常位于被称为转座子 (transposons) 的可移动遗传元件内,因此多重抗性质粒得以进化。R因子通常不会整合到宿主染色体中。
R因子是公共卫生官员关注的主要问题,因为它们可以在细胞群体中迅速传播。这有几个原因。其中之一是许多R因子本身就是接合性质粒。然而,如果一个非接合性的R因子存在于一个同时含有接合性质粒的细胞中,它也可以传播到其他细胞。在这种细胞中,当接合性质粒转移时,R因子有时也会随之转移——即被“动员”。
更令人担忧的是,一些R因子很容易在不同物种间转移。当人类和其他动物摄入抗生素时,带有R因子的宿主细菌的生长会得到促进。然后,这些R因子可以转移到沙门氏菌 (Salmonella) 或志贺氏菌 (Shigella) 等更具致病性的属,从而引发更严重的公共卫生问题。
3、细菌素编码质粒 (bacteriocin-encoding plasmids)
细菌素编码质粒可能为其宿主细菌在微生物世界中提供竞争优势。细菌素 (Bacteriocins) 是能杀死其他细菌的细菌蛋白质。它们通常只对亲缘关系密切的菌株起作用。一些细菌素通过在细胞质膜上形成通道来杀死细胞,从而破坏细胞存活所必需的关键选择性通透性。它们也可能降解DNA和RNA,或攻击肽聚糖 (peptidoglycan) 并削弱细胞壁。
Col质粒 (Col plasmids) 含有合成细菌素(称为大肠杆菌素 (colicins))的基因,这些细菌素针对大肠杆菌。其他质粒则携带针对其他物种的细菌素基因。例如,cloacins能杀死肠杆菌属 (Enterobacter) 物种。一些Col质粒是接合性的,并携带抗性基因。需要注意的是,并非所有的细菌素基因都在质粒上。例如,铜绿假单胞菌 (Pseudomonas aeruginosa) 的细菌素基因(编码称为绿脓菌素 pyocins 的蛋白质)位于染色体上。人体(和其他动物)正常菌群产生的细菌素也是我们抵御入侵病原体的防御机制的一部分。
4、毒力质粒 (virulence plasmids)
毒力质粒编码使其宿主更具致病性的因子。例如,产肠毒素型大肠杆菌 (E. coli) 菌株会引起旅行者腹泻,因为它们含有一个编码肠毒素的质粒。
5、代谢质粒 (metabolic plasmids)
代谢质粒携带编码降解芳香族化合物(如甲苯)、杀虫剂(如2,4-二氯苯氧乙酸)和糖类(如乳糖)等物质的酶的基因。代谢质粒甚至携带了某些根瘤菌 (Rhizobium) 菌株诱导豆科植物结瘤并进行固氮所需的基因。
参考文献
《Microbiology (Seventh Edition)》 | 微生物学,第七版
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-04-30
编制人:叶凡
审稿人:小藻