从基础开始学习原核细胞02:原核细胞膜
来源:武汉市灰藻生物科技有限公司 浏览量:176 发布时间:2026-04-29 14:54:27
1、引言
所有生物都必须有细胞膜。细胞需要与环境进行选择性互动,无论这环境是多细胞生物的内部环境,还是变化更大的外部环境。细胞不仅要获取营养、排出废物,还要在环境变化时,维持内部稳定性。
原核和真核细胞的细胞质都被质膜(plasma membrane)包裹。质膜是细胞与环境接触的主要界面,因此决定了细胞与外界的大部分关系。
原核细胞的质膜尤其重要,因为它承担着多种关键功能。除了包裹细胞质,它还是一种选择性通透屏障:允许特定离子、分子进出,同时阻止其他物质通过。这样既能防止必要成分泄漏,又能控制其他分子的移动。
许多物质无法自行穿过质膜,因此质膜须在必要时协助其运输,运输系统负责营养吸收、废物排泄和蛋白质分泌等。
原核细胞质膜还是多种关键代谢过程的场所,包括呼吸、光合作用、脂质和细胞壁组分的合成等。此外,质膜上还含有特殊的受体分子,帮助原核生物探测并响应周围化学物质。
要理解膜的这些化学差异和多种功能,必须先了解膜的结构。我们先讨论所有膜共有的基本结构,再探讨细菌膜与古菌膜(archaeal membranes)之间的差异。
2、膜结构的流动镶嵌模型(The Fluid Mosaic Model of Membrane Structure)
目前公认的膜结构模型是辛格(Singer)和尼科尔森(Nicholson)提出的流动镶嵌模型(fluid mosaic model)。该模型认为,膜是由脂质双层构成的,蛋白质像“冰山”一样漂浮其中。
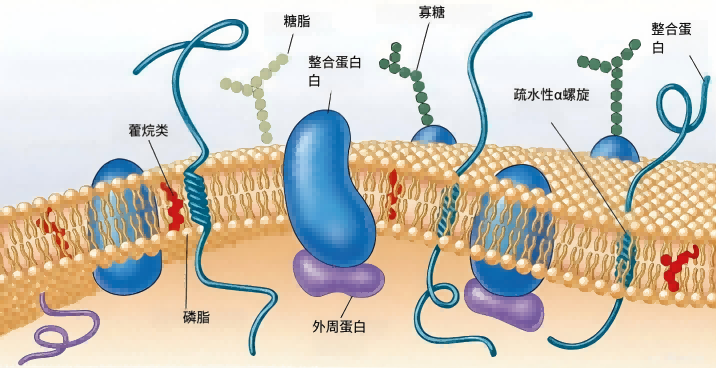
图1 细菌质膜结构
细胞膜经染色后用透射电子显微镜观察,可见细胞膜是非常薄的结构,约5至10纳米厚,呈现为两条深色线条夹着一个未染色的内部区域。这种特征外观表明,膜脂质以两层分子尾对尾的方式排列。
使用冷冻蚀刻技术裂解膜时,可沿脂质双层中心将其分开,暴露出复杂的内部结构。在脂质双层内可见小的球状颗粒,推测是嵌在脂质中的膜蛋白。
膜脂质的化学性质对其形成双层的能力至关重要。大多数膜相关脂质在结构上是不对称的,一端亲水,一端疏水(图2),这类分子被称为两亲性(amphipathic)分子。
亲水端与疏水相互作用,疏水端不溶于水,倾向于彼此聚集。在水环境中,两亲性脂质会自发形成双层。双层膜的外表面是亲水的,而疏水端则埋在内部,远离周围的水。
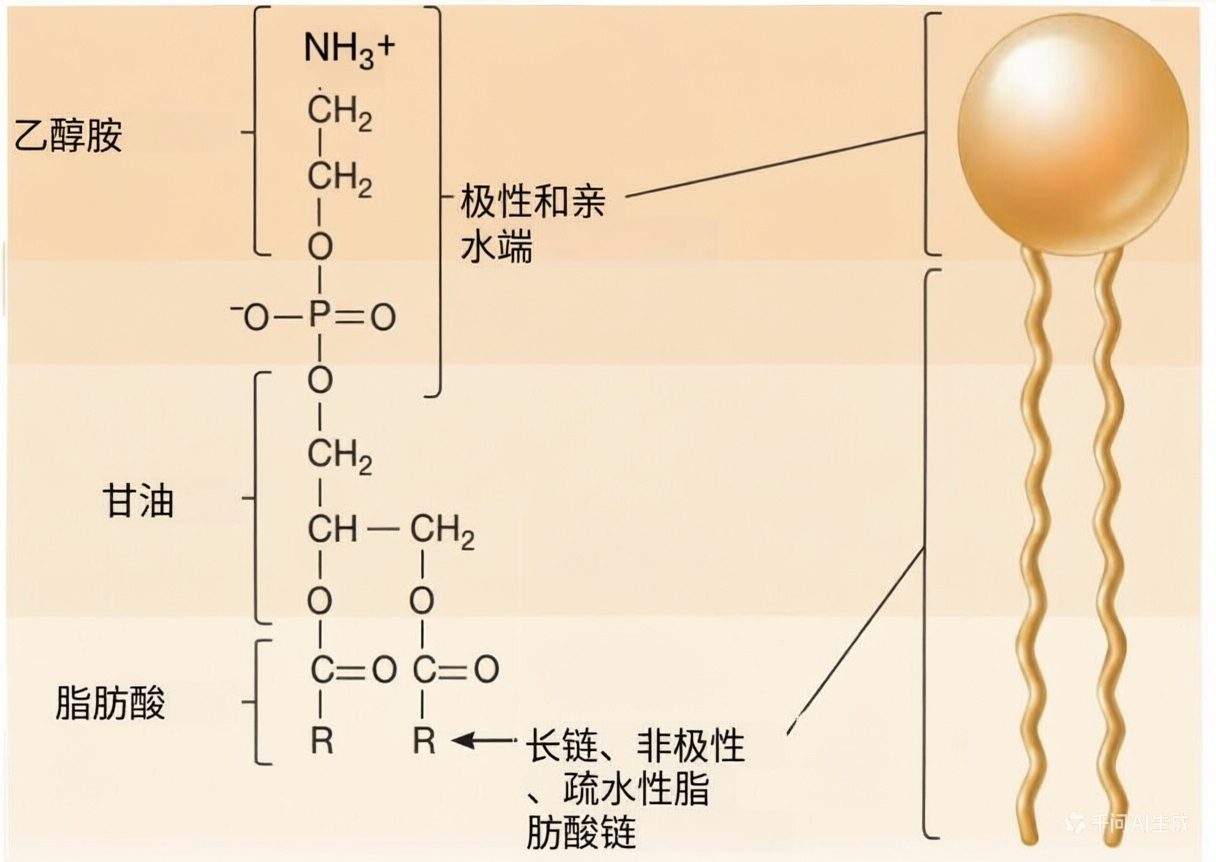
图2 膜脂质的结构
根据与膜分离的难易程度,可将膜蛋白分为两类。
外周蛋白(Peripheral proteins)约占膜蛋白总量的20%到30%,与膜连接松散,易于去除,可溶于水。
整合蛋白(Integral proteins)约占膜蛋白总量的70%到80%,不易从膜上提取,一旦脱离脂质便不溶于水。
整合蛋白和膜脂质一样,也是两亲性的,其疏水区域埋在脂质内部,亲水部分则从膜表面伸出。
整合蛋白可以在膜内横向扩散到新位置,但不会在脂质层中翻转或旋转。碳水化合物常附着在质膜蛋白的外表面,具有重要功能。
3、细菌膜(Bacterial Membranes)
细菌膜与真核膜相似,其许多两亲性脂质都是磷脂。但细菌膜通常不含固醇(sterols),例如胆固醇(cholesterol)(图3),这是与真核膜的主要区别。
但许多细菌膜含有类似固醇的分子,称为藿烷类(hopanoids)(图3)。藿烷类与固醇由相同的前体合成,像真核膜中的固醇一样,起到稳定膜的作用。
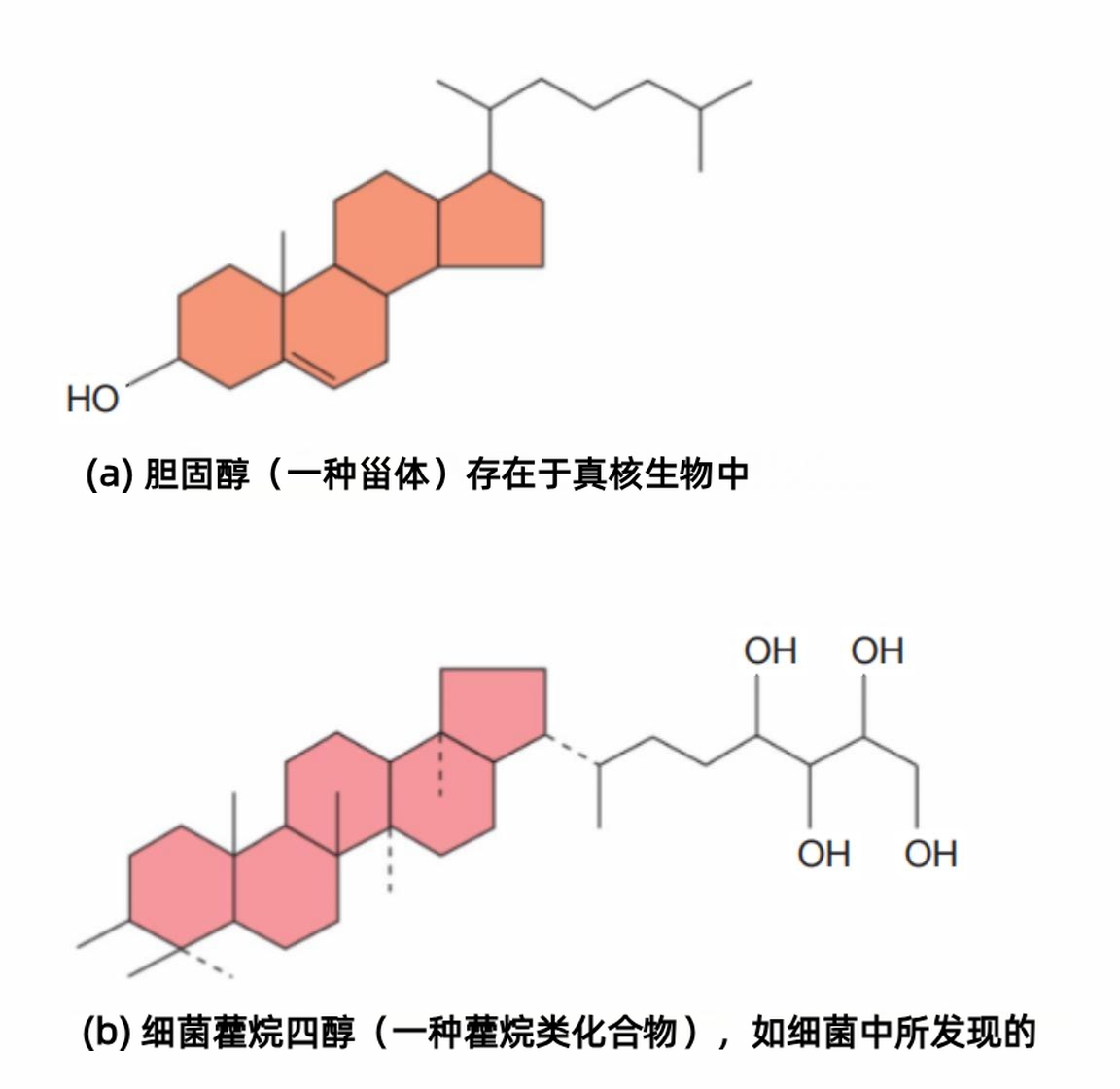
图3 细胞膜的固醇与藿烷类
细菌质膜是一个高度有序、不对称,同时又灵活动态的系统。脂质在质膜中并非均匀分布,而是集中在特定区域。
细菌膜的脂质组成会随环境温度变化,以保证膜在生长过程中保持流动性。例如,在低温下生长的细菌,其膜磷脂中的脂肪酸熔点较低。
尽管原核生物不含线粒体或叶绿体等复杂的膜性细胞器,但在一些细菌中可以观察到内部膜结构(图4)。
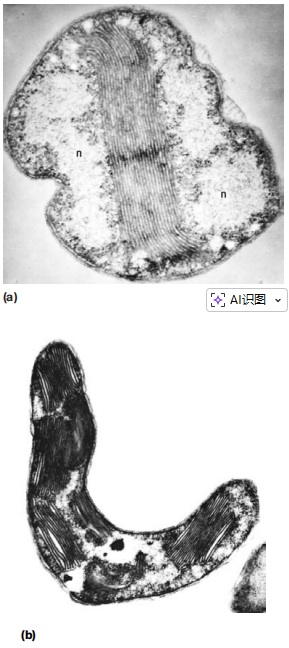
图4 细菌内部膜,a硝化细菌、b光合细菌
质膜内陷在许多细菌中很常见。在蓝细菌、紫色细菌等光合细菌,或亚硝化单胞菌等高呼吸活性细菌中,这种内陷会变得非常复杂。
这些内部膜结构可能是球形、扁平囊泡或管状膜的聚集体。其功能可能是提供更大的膜表面积,以支持更高的代谢活性。
4、古菌膜(Archaeal Membranes)
古菌(Archaea)最显著的特征之一就是其膜脂质的性质。古菌与细菌(Bacteria)和真核生物(Eucarya)不同,其甘油上连接的是通过醚键(ether links)结合的支链烃,而非通过酯键(ester links)连接的脂肪酸(图5)。
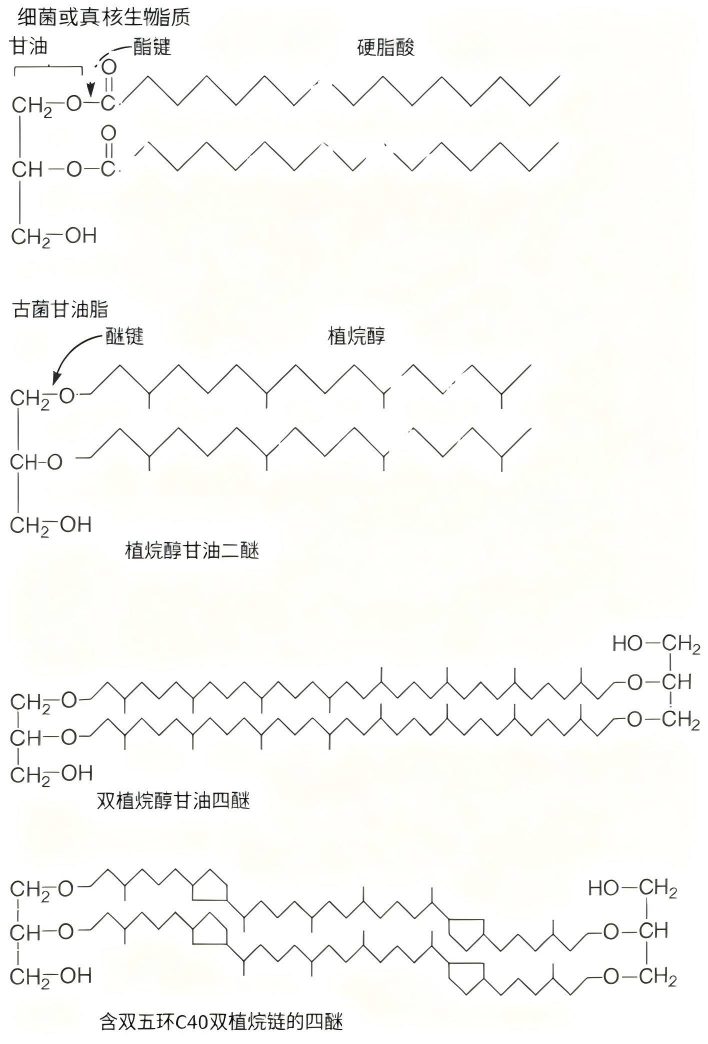
图5 古菌脂质与细菌脂质的区别
有时,两个甘油基团会连接起来,形成极长的四醚(tetraether)。通常,二醚(diether)的烃链含20个碳,四醚的烃链含40个碳。细胞可以通过将烃链环化形成五环结构,来调节四醚的整体长度(图5)。
磷酸基、硫基和糖基可连接到二醚和四醚的第三个碳上,使其成为极性脂质。这些极性脂质在膜中占主导地位,占膜脂总量的70%到93%。
其余脂质为非极性,通常是鲨烯(squalene)的衍生物(图6)。
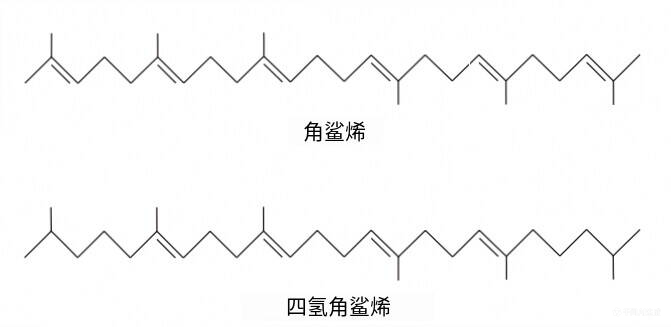
图6 古菌的非极性脂质
尽管膜脂质存在这些显著差异,古菌膜的基本设计仍与细菌和真核生物相似——都有两个亲水表面和一个疏水核心。
当使用C20二醚时,会形成规则的双层膜(图7)。当膜由C40四醚构成时,则会形成刚性更强的单层膜(图7)。
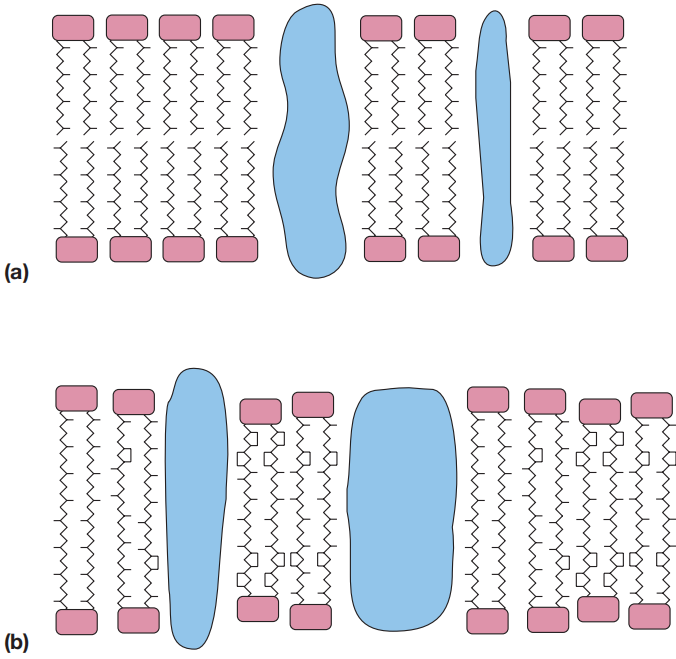
图7 古菌膜示例。(a) 由整合蛋白和C20二醚双层构成的膜。(b) 由整合蛋白和C40四醚构成的刚性单层膜
正如其对稳定性的需求所预期的那样,那些最适生长温度超过85°C的极端嗜热古菌(如热原体 Thermoplasma 和硫化叶菌 Sulfolobus),其膜几乎完全是四醚单层。
生活在中度高温环境中的古菌,其膜则是混合型的,部分区域为单层,部分区域为双层。
下一节我们将详细的介绍细胞质基质。
参考文献
《Microbiology (Seventh Edition)》 | 微生物学,第七版
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-04-29
编制人:叶凡
审稿人:小藻