ATCC真菌处理指南01_培养篇
来源:武汉市灰藻生物科技有限公司 浏览量:2107 发布时间:2024-11-21 00:26:09
ATCC真菌入门指南
ATCC真菌通常以干冰、塑料或玻璃冻存管、玻璃安瓿或血清瓶中的冻干培养物,或培养物形式运输。收到冷冻细胞后,应立即解冻并将其转移到适当的生长培养基中。如果无法立即操作,可将冷冻管存放在液氮蒸气中(温度低于-130°C)。
作为替代方案,冷冻材料可以在-70°C至-80°C的条件下短期储存(1至5天),但温度高于-130°C时,存活率会下降。冻干培养物可在4°C或更低温度下安全储存。收到后,使用无菌蒸馏水或适当培养基重新水化或稀释冻干培养物。
活培养物应立即转移到含有推荐培养基的试管或平板中,并在适当的生长条件下培养。不要将活培养物储存在冷冻室中,因为这可能导致培养物死亡。
提前准备培养基,确保最佳生长条件。

ATCC真菌指南
安瓿打管说明
所有培养物均应被视为潜在危险物,应由受过微生物技术培训的人员,在符合培养物生物安全级别的设施中打开。ATCC建议在生物安全柜中进行玻璃安瓿的处理或打开。如果无法做到这一点,请穿戴防护服、手套、面罩或护目镜,并将安瓿远离身体。确保所有空安瓿在丢弃前已灭菌。
A. 双层安瓿
将安瓿尖端在火焰中加热。
滴几滴水,使玻璃裂开。
用锉刀或铅笔敲碎安瓿。
用无菌镊子取下绝缘层和内管,提起棉塞。
A. 单层安瓿
无菌锉刀在安瓿颈部划痕。
70%乙醇消毒安瓿外表面。
用无菌毛巾或纱布包裹安瓿,以干燥残留乙醇。
将安瓿直立握住并折断开口。确保纱布不要太湿,以免在真空破坏时,酒精被吸入水化干粉。
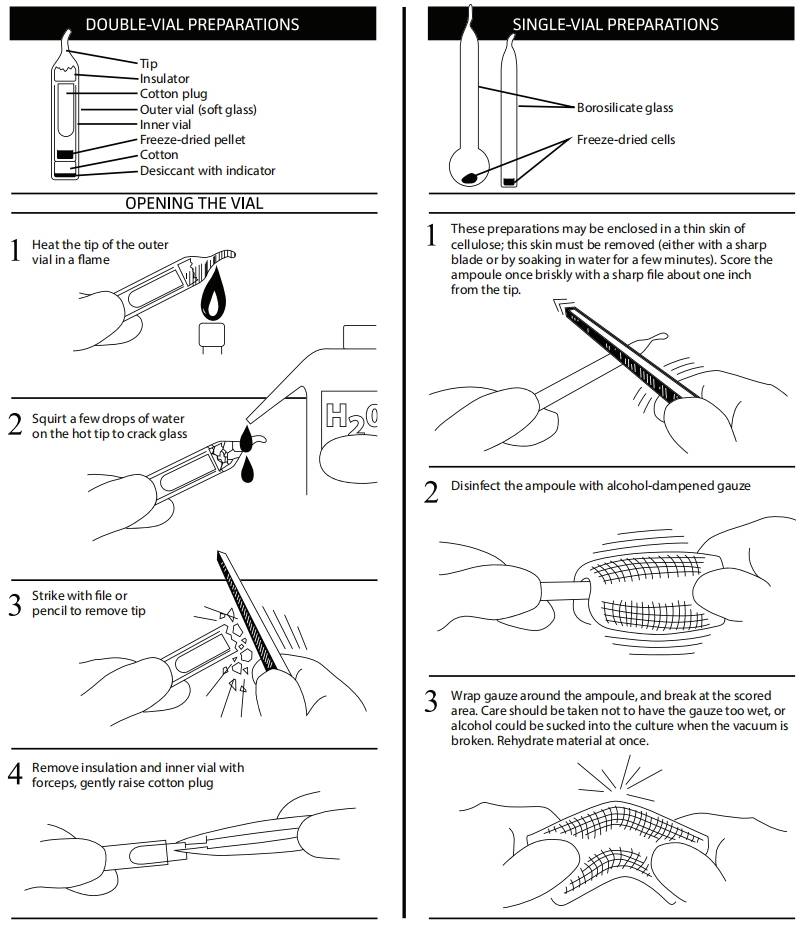
打管说明
冻存物复苏
准备培养基
准备一个含有推荐培养基的无菌试管。确保培养基包含必要成分,并且控制好温度和pH值。
解冻样本
在25°C至30°C的水浴中轻轻摇动样本管,使其迅速解冻;大约需要5分钟,直到所有冰晶完全融化。
消毒处理
将样本管从水浴中取出,并使用新配制的70%乙醇对表面消毒,严格遵守无菌操作条件进行后续操作。
转移培养物
拧开管盖,全部转移到含有适当生长培养基的无菌试管或平板。对于悬浮的酵母细胞,可以将培养物均匀涂布在培养基上以促进单个菌落的生长。注意,许多冷冻的ATCC真菌菌株包含完全生长的培养物制成的琼脂块。
培养
根据推荐条件培养。
检查培养效果
根据推荐的培养期检查培养物,培养周期因菌株而异,具体时间见产品说明
注释:冻存或冻干复苏的某些菌株,可能会表现出更久的培养周期。
冻干粉复苏
加入无菌水
使用巴斯吸管,在无菌条件下向冻干粉加入0.5至0.9毫升无菌水,充分混合。
转移悬液
将全部悬液转移到,含有6毫升无菌水的试管中。
浸泡水化
转移到固体培养基前,让培养物浸泡50至60分钟。对于某些菌株,建议延长水化时间。
培养
根据推荐温度和气体条件培养,可将0.5毫升悬液,转移到其他培养基中来接种。
检查培养物
根据推荐的培养期,检查培养物。
真菌分类介绍
目前,真菌界约有150万种物种,包括各种酵母、锈菌、黑粉菌、霉菌、蘑菇等。这些真核生物在全球范围内广泛分布,涵盖了形态、繁殖和生态多样的大量群体。以下我们将讨论一些已知真菌门的显著特征(表1)。
表1: 已知真菌分类
| 门 | 常见种类 | 区分特征 |
|---|---|---|
| 壶菌门 (Chytridiomycota) | 壶菌 (Chytrids) | 具有单个后鞭毛的动孢子,包含合子减数分裂 |
| 囊菌门 (Glomeromycota) | 丛枝菌根真菌、Glomales、Glomerales | 形成丛枝结构 |
| 接合菌门 (Zygomycota) | 糖霉、粉孢霉 (Sugar Mold, Pin Mold) | 产生厚壁休眠孢子(接合孢子) |
| 芽囊菌门 (Blastocladiomycota) | 芽囊菌 (Blastocladialeans) | 产生动孢子,包含孢子减数分裂。 |
| 子囊菌门 (Ascomycota) | 子囊菌、酵母 (Ascomycetes, Sac Fungi) | 形成子囊 |
| 担子菌门 (Basidiomycota) | 担子菌、伞菌 (Basidiomycetes, Club Fungi) | 产生有性孢子,称为担孢子 |
| 新毛壳菌门 (Neocallimastigomycota) | / | 产生动孢子,存在氢体而非线粒体。 |
| 微孢子虫门 (Microsporidia) | / | 单细胞寄生虫,存在微体而非线粒体。 |
| 卵菌门 (Oomycota) | 卵菌、水霉 (Oomycetes, Water Mold) | 产生大型卵囊 |
壶菌门 (Chytridiomycota)
壶菌门包含多个属,如Spizellomyces、Chytriomyces和Rhizophydium(表2)。壶菌是最小和最简单的真菌,其单细胞或菌丝体由几丁质细胞组成。它们的显著特征是具有单个后鞭毛的动孢子。
像许多真菌物种一样,壶菌主要表现为腐生营养,能够降解几丁质和角蛋白。然而,已有几种物种被发现对植物、动物和其他真菌物种具有致病性。壶菌门的菌株通常栖息于淡水生态系统中,这有助于其生活周期中无性和有性阶段的运动过程。壶菌的生活周期能够在世代之间交替出现单倍体和二倍体阶段,有性繁殖以单倍体为主,无性繁殖以二倍体为主。在有性繁殖过程中,单倍体菌丝体形成雄性和雌性配子囊,从中释放出具有鞭毛的游动配子,随后融合形成二倍体合子。合子萌发后,会产生二倍体菌丝体,其中包含两种类型的孢子囊:一种是释放二倍体动孢子的动孢子囊,用于无性繁殖;另一种是由合子减数分裂产生的孢子囊,释放单倍体动孢子,形成有性繁殖的单倍体菌丝体。
表2: 壶菌门 (Chytridiomycota) 举例
| ATCC编号 | 名称 | 应用 |
|---|---|---|
| 60989™ | Allochytridium luteum Barr 463 | 模式菌株 |
| 28165™ | Chytriomyces hyalinus SOA 1 | 推荐用于课堂演示 |
| 90967™ | Entophlyctis luteolus L-129 | 模式菌株 |
| MYA-2894™ | Monoblepharella sp. M# 15 [JEL 485] | 测序线粒体基因组 |
| MYA-4069™ | Rhizophydium globosum JEL 222 | 模式菌株 |
| 18786™ | Rhizophydium sphaerotheca 70 [IMI 143633] | 推荐用于课堂演示 |
| MYA-1381™ | Aurantioochytrium limacinum SR21 [IFO 32693] | 模式菌株 |
| 18671™ | Spizellomyces plurigibbosus 33 [CBS 668.73, IMI 143638] | 模式菌株 |
| 48900™ | Spizellomyces punctatus 117 [NG-3] | 模式菌株,基因组测序菌株 |
| 18785™ | Triparticalcar arcticum 59 [CBS 666.73, IMI 143636] | 模式菌株 |
囊菌门 (Glomeromycota)
囊菌门是全球陆地生态系统中的生物营养体,它们与当地植物根系形成独特的共生关系。形成共核菌丝,通过无性繁殖形成孢子。在适宜条件下,孢子萌发,与植物根系形成丛枝结构,建立新的菌根共生关系。囊菌门的代表属包括Geosiphon、Glomus和Acaulospora。
在共生关系中,囊菌门与陆地植物的根系或菌丝体形成丛枝菌根,以获取碳和能量。然而,有证据表明,囊菌门可能独立存在,不形成共生关系。
玉米黑粉菌
接合菌门 (Zygomycota)
接合菌门包含多个属,如Conidiobolus、Erynia和Mucor(表3)。近年来,传统上归类为接合菌门的真菌进行了系统分类重组。接合菌门包括破坏性的糖霉或针霉,但该门内的许多物种可以与植物形成共生关系。接合菌门通常以菌丝或丝状体的形式生长,每种菌丝或丝状体均由几丁质-壳聚糖细胞壁组成。
该陆地门的成员能够进行有性和无性繁殖,有性繁殖以单倍体为主,无性繁殖以二倍体为主。在无性繁殖过程中,无性孢子(孢囊孢子)在孢子囊中产生。孢囊孢子萌发后,形成体菌丝结构。在有性繁殖过程中,不同的交配菌株(原配子囊)相互生长并融合形成合子。合子随后发育形成接合孢子,这是接合菌门的特征厚壁休眠孢子。接合孢子会在一段时间的休眠后重新开始繁殖周期。
表3: 接合菌门 (Zygomycota) 举例
| ATCC® 编号 | 生物命名 | 应用 |
|---|---|---|
| 24418™ | Conidiobolus obscurus [CBS 182.60] | 模式菌株 |
| 32803™ | Entomophthora destruens NRRL 3727 [CBS 208.65] | 模式菌株 |
| 34698™ | Erynia blunckii Nr. 2 [CBS 194.77, IMI 211910] | 模式菌株 |
| 208867™ | Eryniopsis ptychopterae ARSEF 2671 [KVL48] | N/A |
| MYA-4580™ | Furculomyces boomerangus AUS-42-7 | 同型生殖的Harpellales |
| 36965™ | Mortierella alpina 84 | 降解造纸厂废物 |
| MYA-4767™ | Mucor ellipsoideus UTHSC 02-2090 [FMR 10021, CBS 126271] | 模式菌株 |
| MYA-4621™ | Rhizopus oryzae FGSC 9543 [RA99-880] | 基因组测序菌株(美国Broad研究所) |
| MYA-4582™ | Smittium angustum AUS-126-30 | 毛霉目肠道真菌 |
| 208865™ | Zoophthora radicans ARSEF 4784 [KVL610] | N/A |
芽囊菌门 (Blastocladiomycota)
芽囊菌门目前被认为是壶菌门和新毛壳菌门的“姊妹门”。最初,芽囊菌类真菌被归类为壶菌门,但最近的DNA分析将其指定为一个独立的门。芽囊菌门是栖息于淡水或土壤中的动孢子真菌。类似于壶菌门,芽囊菌门物种要么是腐生的,要么寄生于藻类、陆地植物、真菌和无脊椎动物。
芽囊菌门与其姊妹门壶菌门在生活周期上有所不同。与壶菌门利用合子减数分裂不同,芽囊菌门利用孢子减数分裂,这意味着合子通过有丝分裂产生多细胞二倍体孢子体。这个孢子体通过减数分裂产生孢子,这些孢子再通过有丝分裂产生单倍体个体,称为配子体。目前的芽囊菌门代表属包括Allomyces、Blastocladia和Coelomomyces。
表4: 芽囊菌门 (Blastocladiomycota) 示例
| ATCC® 编号 | 生物命名 | 应用 |
|---|---|---|
| 10984™ | Allomyces javanicus [CBS 600.78] | N/A |
| 38327™ | Allomyces macrogynus Burma 3-35 | 基因组测序菌株和课堂演示 |
| 44886™ | Allomyces macrogynus asn 1 | ATCC® 38327的营养缺陷型突变体 |
子囊菌门 (Ascomycota)
子囊菌门的成员通常被称为囊菌,因为它们的定义特征是子囊(ascus),子囊内含有通过自由细胞形成而不具运动能力的子囊孢子,形似囊状。这种结构用于子囊菌门生活周期的有性生殖阶段。子囊孢子萌发后,形成体菌丝,随后发展成产孢结构——分生孢子器。分生孢子器是子囊菌门生活周期中无性、主导阶段的一部分,负责产生和快速传播无性生殖孢子,即分生孢子。
该门内的许多物种以丝状菌丝体的形式生长,细胞壁中含有几丁质和β-葡聚糖。然而,子囊菌门中的某些物种,如念珠菌属(Candida)和酿酒酵母属(Saccharomyces),通常以酵母形式生长(表5)。这些后者通过出芽方式进行繁殖。子囊菌门成员对人类特别重要,因为它们是抗生素的来源,并用于面包、奶酪、啤酒和葡萄酒等食品的生产。然而,该门的其他成员是多种物种的病原体。例如,子囊菌门中的植物病原体可引起苹果黑星病、稻瘟病和荷兰榆树病。

疣状瓶霉
表5: 子囊菌门 (Ascomycota) 示例
| ATCC® 编号 | 生物命名 | 应用 |
|---|---|---|
| 10231™ | Candida albicans 3147 [CBS 6431, CCY 29-3-106, CIP 48.72, DSM 1386, IFO 1594, NCPF 3179, NCYC 1363, NIH 3147, VTT C-85161] | 多种应用的参考菌株 |
| 6205™ | Chaetomium globosum QM 459 [1042.4, CBS 148.51, CBS 161.52, CEB 1218.1, CEB 1218.2, CECT 2701, DSM 1962, IFO 6347, IMI 45550, MUCL 1984, NRRL 1870, UPSC 3159, VTT D-81079] | 各种测试的参考菌株 |
| 76902™ | Dipodascopsis tothii CBS 759.85 [CSIR 31, NRRL Y-12690] | 在子囊孢子发生期间产生前列腺素 |
| 26050™ | Gerlachia nivalis | 雪霉,对黑麦或草根有致病性 |
| MYA-4619™ | Neurospora crassa FGSC 9013 [OR74A, N10] | 基因组菌株和遗传学研究 |
| 26475™ | Neurospora crassa 74A-T28-M1 | 遗传学研究 |
| 204508™ | Saccharomyces cerevisiae S288C | 基因组测序菌株,许多遗传菌株的亲本菌株 |
| 24843™ | Schizosaccharomyces pombe HFBDG41 | 基因组菌株和遗传模型菌株 |
| 13631™ | Trichoderma reesei QM 6a [CBS 383.78, IMI 192654, IMI 45548, T.V. B117] | 酶生产,参考菌株,基因组菌株 |
| 18377™ | Xanthoria parietina | 地衣共生体 |
担子菌门 (Basidiomycota)
担子菌门是一组多样化的真菌,通常被称为担子菌,包括蘑菇、牛肝菌、马勃、地星、臭菌、鸟巢菌、胶质菌、层孔菌、锈菌和黑粉菌等。担子菌门的代表属包括Agaricus、Pleurotus和Ganoderma(表6)。这些丝状真菌的主要特征是产生有性孢子,称为担孢子,这些孢子位于特化的孢子产生结构——担子的外部。
在大多数担子菌中,除了锈菌和黑粉菌外,担子是在称为担子果的子实体中产生的。担子菌门的有性生殖最终产生担子果中的担孢子。相比之下,担子菌的无性生殖通过菌丝断裂、分生孢子、节孢子或卵孢子的产生来进行。后者是某些担子菌,包括许多酵母,唯一的繁殖方式。
担子菌广泛存在于大多数陆地生态系统、淡水和海洋环境中。这些真菌帮助分解死亡的有机物质,因此在维持其生态系统中起着重要作用。担子菌在人类事务中也非常重要,一些物种作为病原体或本身有毒,而另一些物种则可用作食物来源。例如,黑粉菌是谷物和作物的病原体,已被发现对大麦、小麦、燕麦和玉米等禾本科作物造成毁灭性影响。相反,许多蘑菇种类,如香菇和杏鲍菇,常被食用。
表6: 担子菌门 (Basidiomycota) 示例
| ATCC® 编号 | 生物命名 | 应用 |
|---|---|---|
| 36981 | Agaricus bisporus D621 | 商业白蘑菇 |
| 34620 | Boletus edulis CBS 455.69 | 食用蘑菇 |
| MYA-4618 | Coprinopsis cinerea FGSC 9003 [okayama7#130] | 蘑菇,基因组测序菌株 |
| 200597 | Ganoderma lucidum | N/A |
| MYA-4612 | Malassezia globosa CBS 7966 [ATCC 96807, GM 35] | 基因组菌株,人类机会性病原体 |
| MYA-4611 | Malassezia restricta CBS 7877 [ATCC 96810, JCM 14890, RA 42.2C, NBRC 103918] | 基因组菌株,人类机会性病原体 |
| MYA-4764 | Phanerochaete chrysosporium 2-2786 | Hite腐朽菌,基因组测序菌株 |
| 96999 | Pleurotus ostreatus CCB 004 | 灰平菇 |
| MYA-4550 | Sporobolomyces salmonicolor VITEK 301525 [MUCL 51628] | VITEK2的参考菌株 |
| 20509 | Cutaneotrichosporon oleaginosus D | 通过纤维素降解生产油 |
新毛壳菌门 (Neocallimastigomycota)
新毛壳菌门目前被认为是芽囊菌门和壶菌门的“姊妹门”。该门由存在于许多大型哺乳动物反刍胃和后肠中的厌氧真菌组成。新毛壳菌门的物种在其哺乳动物宿主中消化纤维方面发挥着关键作用,产生用于分解植物材料的多糖降解酶。新毛壳菌门与姊妹门类似,也产生具鞭毛的动孢子。
然而,这些动孢子缺乏大多数壶菌中存在的非鞭毛中心粒。此外,新毛壳菌门还缺乏线粒体,而是拥有一个称为氢体的细胞器,该细胞器在厌氧条件下产生分子氢作为能量生成的副产物。新毛壳菌门的代表包括Orpinomyces、Piromyces、Neocallimastix、Caecomyces和Anaeromyces。
微孢子虫门 (Microsporidia)
微孢子虫门由形成孢子的单细胞寄生虫组成。该门的一些例子包括Enterocytozoon、Pleistophora和Encephalitozoon等属。这些真菌生物曾被认为属于原生动物,主要感染节肢动物和鱼类。然而,微孢子虫由于能够感染免疫受损个体而在医学上具有重要意义。这些生物的独特之处在于,它们拥有真核生物中最小的基因组之一。此外,微孢子虫缺乏线粒体,而是拥有线粒体的简化形式——微体。
微孢子虫在宿主体内肠道系统中复制。摄入的微孢子虫孢子萌发并膨胀至破裂,迫使内部的细丝迅速射出感染性内容物,称为孢浆,进入周围的宿主细胞。一旦进入宿主体内,孢浆生长形成微孢子虫生物;成熟后,该生物产生新的孢子,通过粪便污染和吸入传播到新的宿主。这种生活周期在不同物种之间差异很大,包括无性和有性繁殖的不同形式。
卵菌门 (Oomycota)
卵菌门的成员通常被称为水霉,是一种假真菌、致病性真核生物。卵菌门的代表属包括Phytophthora、Pythium和Saprolegnia(表7)。这些生物广泛存于全球,可导致植物疾病,如马铃薯晚疫病和橡树猝死病。卵菌门的形态可以是单细胞或丝状分枝,后者很少含有隔膜。卵菌的细胞壁结构由纤维素和β-1,3-及β-1,6-葡聚糖组成。
卵菌既进行有性生殖也进行无性生殖。卵菌的有性生殖通常是异型配子囊的,大多数物种的配子囊分化成小的菌丝状雄性结构,称为雄器,和较大的球形雌性结构,称为藏卵器。这些有性结构可以由不同的菌丝体或同一菌丝体发育而来,但同一菌丝体上产生的配子囊可能不相容。
在有性生殖过程中,发育中的雄器被激素吸引到藏卵器,从而产生受精管。来自雄器的减数分裂产生的单倍体核通过受精管引入到卵球中,与卵球核融合。受精的卵球随后在藏卵器内发育成卵孢子,成熟后释放。卵孢子萌发后,发育成二倍体菌丝体。
在无性生殖过程中,大多数卵菌通过异形动孢子进行繁殖。根据卵菌的种类,这些孢子会发育成孢子囊或从孢子囊突出的囊泡。在许多物种中,会产生两种形态上不同的动孢子:初生动孢子和次生动孢子。初生动孢子呈梨形,鞭毛附着在孢子的前端。次生动孢子,由大多数卵菌产生,呈肾形,鞭毛侧向插入孢子表面的凹陷处。
表7: 卵菌实例
| ATCC® 编号 | 生物名称 | 应用 |
|---|---|---|
| MYA-4127™ | Phytophthora infestans T30-4 | 基因组测序菌株 |
| MYA-4040™ | Phytophthora megakarya p42 [22H7, 203532] | 对可可树具有致病性 |
| MYA-2949™ | Phytophthora ramorum Pr-102 | 基因组测序菌株 |
| MYA-4756™ | Phytophthora sojae p236 [P6497, 28F9] | 基因组测序菌株 |
| 66260™ | Pythium carolinianum 85-54 | 对樱桃种子具有致病性 |
| 28251™ | Pythium insidiosum K169-B91 [A-25710] | 对马具有致病性 |
| 201957™ | Pythium oligandrum 91-4 | 潜在的根部病原体生物控制剂 |
| 36145™ | Saprolegnia diclina P203 | 寄生于鲈鱼 |
| MYA-399_TT™ | Saprolegnia salmonis NJM 9851 | 对鲑鱼具有致病性 |
| 38488™ | Saprolegnia shikotsuensis SANK 23 177 | 鱼类寄生虫 |
真菌的生长条件
营养需求
真菌界主要由异养腐生生物组成,通过吸收有机物质作为营养来源生存。为了消耗有机物质,真菌必须释放消化酶,将大的碳水化合物、蛋白质和脂类分解成较小的可利用分子,这些分子可以轻松地穿过细胞壁和细胞膜被吸收。真菌能否在环境中生存很大程度上取决于其产生的消化酶类型。最终,为了使这一过程成为可能,真菌物种必须生长在一个有水源的地方。
相当多的真菌寄生在植物、动物或其他真菌上生存。这些物种中的许多仍然能够过腐生生活方式,而另一些则是专性寄生生物。专性寄生的真菌物种在营养需求上相对受限,通常需要特定物种才能生存。
离蠕孢霉
环境条件
真菌的生长很大程度上依赖于环境的湿度、温度、pH值和大气条件。如下所述,这些要求在不同物种间可能存在显著差异。
湿度
所有真菌物种都需要水分存在;然而,所需水量和渗透性有所差异。有些只需要少量水分以助于吸收,而其他物种则能在高盐含量的基质上生存。
温度
大多数真菌物种可以在10°C至40°C的温度范围内生存,最佳温度在25°C至30°C之间。其他极端嗜好者物种能够在这些温度范围之外的热嗜好或冷嗜好条件下生存。理想生长温度会因物种而异(参见:注释2)。
pH值
许多真菌在较低的pH水平(4-7)下茁壮成长。然而,这个pH范围可能会影响受代谢过程。如果pH值超出最佳范围,真菌的生长可能会受到影响。
气体条件
通常真菌都是需氧的,然而,也有特殊情况,比如:专性厌氧(如新毛壳菌门)、兼性厌氧(如Trichocladium canadense等)。
特定真菌培养
卵菌属 (Phytophthora spp., 卵菌门)
卵菌属的复苏相对困难,冻存后存活率较低。此外,一些卵菌属物种生长缓慢,对温度敏感,且营养需求挑剔。ATCC复苏程序:
解冻
置于25°C至30°C的水浴中,直至完全解冻(大约5分钟)。将安瓿浸入水中,深度足以覆盖冷冻材料。不要摇晃安瓿。
无菌转移
解冻后立即用新配制的70%乙醇擦拭安瓿,并将冷冻样本无菌转移到一小体积(约5.0 mL)的营养肉汤或酵母霉菌肉汤(YM肉汤)中,其中含有微量的生物素和硫胺素。在推荐温度下培养,直到可见生长;这可能需要长达20天。
转移至湿润的琼脂培养基
然后将培养物转移到湿润的琼脂培养基上,如V8-黑麦培养基(ATCC培养基1970)、V8果汁琼脂(ATCC培养基343)和菜豆培养基(ATCC培养基322)(参见:注释3)。
注释3:为了提高卵菌复苏成功率,建议先在营养肉汤中培养,然后再转移到黑麦、V8或菜豆基培养基上。
白假丝酵母
真菌计数
真菌计数通常用于监测生长速率,或者制作定量培养物。酵母和产孢子的真菌,通常采用菌落数(CFU/mL)来计数。
要计算悬液CFU/mL,从活跃生长的溶液中取出一定量的样品,并进行连续稀释。稀释的程度取决于菌株的生长速率和生长阶段。每个稀释度的样品应均匀涂布在琼脂平板上。经过合适的生长周期后,计数每个平板上的菌落数。为了获得最佳结果,只使用菌落数在25-250之间的平板,这将提供准确的计数表示。例如,如果从一组最终稀释度为10^-7的平板中得到的三个计数分别为40、37和43,则计数为4.0 x 10^8 CFU/mL。
酵母和产孢子真菌的生长速率和培养要求在不同物种间差异很大,因此很难量化计数。ATCC建议快速读取生长产孢子真菌的平板,因为培养物会迅速长在一起,个体菌落将不再可区分。
培养基
真菌培养基通常是蛋白质、微量元素、氨基酸和碳水化合物的混合物。除了提供营养外,培养基还帮助维持pH值。pH值可以通过一种或多种缓冲系统(如MOPS或磷酸钾)来维持。ATCC使用多种类型的培养基,以提供每种真菌菌株的最佳生长条件。
常用培养基
YM琼脂 (ATCC 200) 常用于酵母、霉菌和其他喜酸微生物。由酵母提取物、葡萄糖、麦芽提取物和蛋白胨的混合物制成,为微生物提供微量元素、维生素、氨基酸、碳源和蛋白质。
埃蒙斯改良萨布罗琼脂 (ATCC 28) 用于病原性和共生真菌和酵母培养。由葡萄糖和新蛋白胨按2:1的比例混合而成。这种培养基是萨布罗葡萄糖琼脂的改良版,含有较少的葡萄糖,维持在中性pH值,有助于某些病原性真菌生长。
马铃薯葡萄糖琼脂 (PDA) (ATCC 336) PDA是一种常用的微生物培养基,用于培养植物致病株真菌和细菌物种。通常由过滤的煮沸马铃薯悬浮液与葡萄糖和琼脂混合而成。
酵母提取物蛋白胨葡萄糖 (YEPD) (ATCC 1245) 用于酵母培养。由酵母提取物、蛋白胨和葡萄糖按1:2:2的比例混合而成。这种培养基调整至pH 5.6。
麦芽提取物琼脂 (ATCC 325) 推荐用于酵母和霉菌的检测、分离和计数。这种培养基由麦芽提取物、葡萄糖和蛋白胨的混合物制成。
玉米粉琼脂 (ATCC 307) 用于真菌培养和厚垣孢子产生。玉米粉提供的碳源、氮源和维生素是真菌生长所必需的。
V8果汁琼脂 (ATCC 343) 推荐用于产孢子和各种真菌生命周期特性检查。培养基含有V8果汁,并补充了碳酸钙。
组分介绍
葡萄糖 葡萄糖(D-葡萄糖)
是一种高度营养的补充剂,用于培养挑剔的微生物、老化培养物和接种量小的培养物。可以用作葡萄糖肉汤/琼脂培养基,也可以作为复杂或定义培养基中的补充剂。葡萄糖主要用作碳源。
蛋白胨
蛋白胨是水溶性的蛋白质衍生物,通过酶或酸部分水解动物蛋白质制备的。不是所有的微生物都能利用大气中的氮气。许多物种需要有机或无机固定氮。蛋白胨作为有机氮源常用于无血清培养基。蛋白胨的营养价值取决于其提供的必需氮的氨基酸含量。蛋白胨的起始材料可以是动物或植物来源,包括肉类、大豆、酪蛋白和乳清。
酵母提取物
酵母提取物是从自溶的酿酒酵母细胞中提取。在自溶过程中,内源性酵母消化酶将蛋白质分解成肽和氨基酸,可以被真菌用作氮源。此外,酵母提取物还提供水溶性的B族维生素、碳水化合物和游离谷氨酸。
麦芽提取物
麦芽提取物是麦芽大麦的水溶性部分。它含有高浓度的还原糖。为了生成麦芽提取物,首先让大麦发芽(称为发芽过程)。然后通过磨碎大麦并在温度调节的水中加热大麦来诱导酶的产生和随后的糖分解。最终的物质(“麦汁”)经过过滤和干燥,用于培养基中。
补充添加剂
某些真菌和酵母的生长,可能需要添加基础培养基中不易获得的成分,如盐类和微量元素。添加补充剂后,培养基的保质期应根据具体情况确定。含有补充剂的培养基不应冷冻,因为这可能导致某些化合物沉淀。培养基应储存在2°C至8°C之间。
参考资料
https://www.atcc.org/resources/culture-guides/mycology-culture-guide
【相关资源】
名称:致病疫霉 | Phytophthora infestans
微生物资源鉴定保藏平台
敬请关注“灰藻视界”,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共同创造生命科学的未来!
更新日期:2024-11-20
#创作团队
编制人:小藻 | 审稿人:小灰