酵母与细菌共培养
来源:武汉市灰藻生物科技有限公司 浏览量:77 发布时间:2026-05-22 13:25:17
摘要
酵母与细菌共培养在自然界普遍存在,目前已广泛应用于食品领域(如乳制品、酒精饮料)。其优势包括:拓宽底物范围、提高生产效率、改善产品营养。
目前从平板到生物反应器的工艺已开发完善,但工业化仍面临挑战——许多菌间相互作用尚未阐明。传统离线测量仍是主要手段,在线单细胞监测技术虽带来新可能,但应用仍处起步阶段。
本综述聚焦细菌、酵母及二者共培养的工艺与过程分析最新进展。建立适宜的监测方法,将加速工业共培养工艺开发,推动技术标准化与跨地区应用。
一、引言
在限定环境中,涉及两种或两种以上已知微生物的共培养,已被认为是诱导特定产物生物合成以及调控机制(包括辅因子和代谢物交换)的有效方法。
1.1 纯培养的局限性
传统上,微生物生物生产多采用无菌纯培养或非无菌混合培养。纯培养由于缺乏最佳的生物因素(如细胞间通讯与相互作用)和非生物因素(如温度、pH值、氮源和碳源),其生长状况往往与自然环境中的生长大相径庭。
1.2 共培养的优势
提高产量:每种微生物都能刺激其他微生物,从而提高特定代谢物的产量
增强稳健性:对培养条件变化具有更高的适应能力
缓解代谢负担:通过菌株间转化过程的共享及辅因子交换,有效缓解中间代谢物过量积累的问题
应对复杂原料:更能应对复杂原料组成的挑战
1.3 共培养的局限性
底物竞争导致产品专一性降低
底物吸收无法精确分配给单一微生物
1.4 监测技术的重要性
由于共培养系统比纯培养更难控制,引入能够提供足够时间分辨率和相关数据的监测工具至关重要。监测技术可分为三类:
实时监测:直接在细胞悬浮液中或其附近进行
在线监测:位于培养系统旁边
离线监测:需在一定空间距离外进行,需要样品转移
目前适用的过程分析技术包括:散射光谱、吸光度与自荧光光谱、荧光标记以及流式细胞术。
1.5 本综述的目标
✅ 概述共培养体系合成多种产品(尤其是食品应用)的最新进展
✅ 探讨用于监测和控制微生物群落的适当工具
✅ 重点关注单细胞分析技术及其机遇与挑战
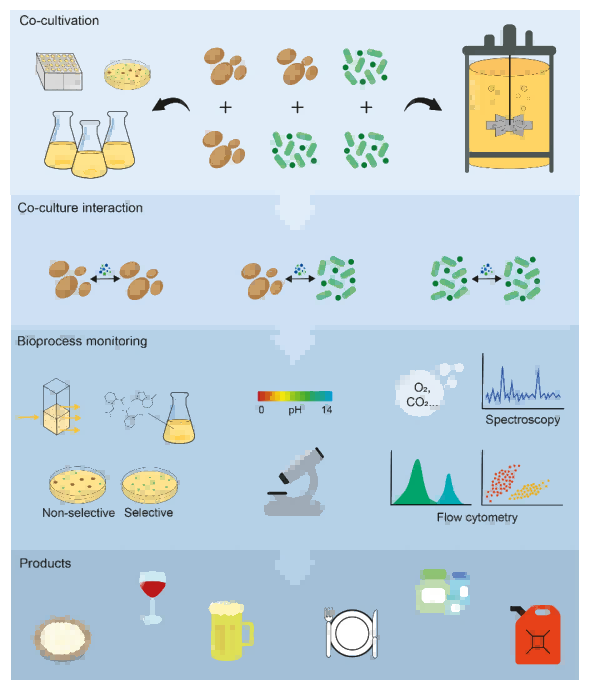
图1. 展示了酵母与细菌共培养的不同培养规模、监测技术及产品应用前景。
二、微生物共培养的类型与应用
自然界存在多种共生关系,如固氮细菌与豆科植物、真菌共生体与植物等。通过模拟这些系统,相关研究可应用于生物技术工艺。
共培养技术已广泛应用于食品开发,涵盖乳制品、酒精饮料及替代蛋白等,用于赋予食品独特的颜色、风味、口感和质地。
(说明:本综述中所描述的共培养比例均以 CFU/mL 为单位,除非另有说明。)
2.1 酵母共培养
(1)葡萄酒酿造
研究案例:酿酒酵母 QA23 与托鲁氏酵母 Zymaflore® alpha 共培养
方法:在含生长因子(麦角甾醇、油酸等)的膜反应器中进行
关键发现:通过调整生长因子浓度,可有效控制两种菌群的比例
(2)啤酒发酵
研究案例:托鲁氏酵母 Biodiva™ 与酿酒酵母 Safale S-04™ 共培养
接种比例效果:
1:2 → 托鲁氏酵母提前死亡
1:20 → 酿酒酵母生长受抑制
风味化合物变化:
癸酸乙酯:较纯培养提高 46.7%~70.1%
十二酸乙酯:较纯培养提高约 148%
(3)乙醇生产
研究案例:固定化酿酒酵母 PE-2 与谢弗氏假丝酵母共培养
方法:细胞固定于海藻酸钙珠
效果:
乙醇产量较纯酿酒酵母培养提高 4 倍
乙醇产量较纯谢弗氏假丝酵母培养提高了3倍
(4)乙酸乙酯生产
研究案例:酿酒酵母 Y3401 与异常威克汉姆酵母 Y3604 共培养
接种方式比较:同时接种 > 顺序接种
最优条件:酿酒酵母:威克汉姆酵母 = 3:1,产量达 6.41 g/L
(5)果寡糖生产
研究案例:出芽短梗霉与酿酒酵母共培养
方法:固定化出芽短梗霉 + 包埋后10小时接种酿酒酵母
效果:果寡糖含量达 131.9 g/L,较对照组提高约 23 倍
(6)药物前体生产
研究案例:热带假丝酵母与酿酒酵母共培养生产苯乙酰卡宾醇
底物:甘蔗渣水解液
效果:产量 8.65 mmol(纯培养仅 4.50 mmol)
优化:两相乳化体系使生产效率提高 1.5 倍
2.2 酵母-细菌共培养
(1)开菲尔发酵
牛奶开菲尔:菌粒由细菌多糖、乳酸菌、乙酸菌和酵母组成,具有心脏保护、抗病毒、抗癌等多种健康益处。
水开菲尔:选定瓦尔比酵母菌和肠膜明串珠菌优化后获得:
乙醇:7 g/L
乳酸:4.98 g/L
(未优化对照组:乙醇 23 g/L,乳酸仅 1.33 g/L)
(2)γ-氨基丁酸生产
菌株:酿酒酵母 SC125 + 粪肠球菌 AB157
效果:产量提高 1.74 倍
机制:与 GABA 合成相关基因(glnH、glnM、gad、gadA)表达上调
(3)胞外多糖生产
菌株:鼠李糖乳杆菌 + 酿酒酵母
效果:产量提高 39%~49%
最佳菌株:RW-9595 M
(4)花生大豆奶发酵
菌株:嗜酸乳球菌 + 嗜酸乳杆菌 + 酿酒酵母
效果:
乳酸浓度:8.51 g/L(单菌培养仅 0.03~3.26 g/L)
碳水化合物消耗率:58%~78%
氨基酸消耗量明显更高
(5)啤酒中乳酸菌存活
问题:啤酒花中的异α-酸具有抗菌性,抑制乳酸菌生长
解决方案:副干酪乳杆菌 L26 + 酿酒酵母 S-04 共培养
效果:乳酸菌活菌数达 10⁹ CFU/100 mL(单菌培养7天后无活菌)
(6)白酒酿造
菌株:酿酒酵母 + 面包乳杆菌
关键因素:
低还原糖(~15 g/L)有利于面包乳杆菌生长(提高1.8倍)
高乙醇(40 g/L)抑制面包乳杆菌(下降1.56倍)
(7)香肠发酵
菌株:汉氏德巴利酵母 + 木糖葡萄球菌 + 戊糖片球菌
比例:4:1:0.2
机制:促进α-酮酸生成,实现氨基酸交叉供给
效果:支链醛类浓度明显高于单菌培养
(8)Nisin 生产
菌株:乳酸乳球菌 + 酿酒酵母
最优比例:100:1(乳酸乳球菌:酿酒酵母)
效果:nisin 产量提高 81.4%
(9)双歧菌素 A 生产
菌株:动物双歧杆菌 + 异常威克汉姆酵母 Y-5
效果:产量提高 3 倍
(10)白藜芦醇生产
菌株:工程大肠杆菌 + 酿酒酵母
机制:大肠杆菌分泌对香豆酸 → 酿酒酵母转化为白藜芦醇
最优条件:1:1 接种,33.5℃
产量:36 mg/L
(11)含氧紫杉烷类生产
菌株:工程酿酒酵母 + 大肠杆菌
关键策略:使用木糖替代葡萄糖,避免乙醇积累抑制大肠杆菌
最终产量:33 mg/L
(12)生物燃料生产
菌株:专性厌氧菌(植物发酵梭菌)+ 工程酿酒酵母
机制:酵母消耗氧气,缓解厌氧菌受到的抑制
效果:
共培养乙醇产量:22 g/L
纯培养:6~9 g/L
菌群稳定维持 50 天
2.3 细菌共培养
(1)微生物蛋白生产
菌株:甲烷氧化菌 + 氢氧化菌
效果:必需氨基酸比单菌培养高出 2.4~6.1 倍
(2)维生素 B12 富集
菌株:弗氏丙酸杆菌 + 解淀粉芽孢杆菌
最优比例:1000:1
条件:厌氧48小时 + 需氧24小时
(3)2-酮-L-古洛糖酸生产
菌株:氧化葡萄糖杆菌 + 普通酮古洛糖酸菌
优化效果:
初始产量:12.9 g/L(~15%)
优化后:59.1 g/L
基因工程改造后:76.6 g/L(达工业水平 89.7%)
(4)ABE 发酵
菌株:纤维梭菌 + 贝耶林克氏梭菌
共生机制:纤维梭菌糖化木质纤维素并生成丁酸 → 贝耶林克氏梭菌利用糖和丁酸生产丁醇
最优条件:同步接种、1:10 比例、前24小时控制 pH
效果:ABE 产量 11.8 g/L,提高 108%
(5)含 nisin 的细菌纤维素
菌株:木醋杆菌 + 乳酸乳球菌
最优比例:1:4(木醋杆菌:乳酸乳球菌)
效果:nisin 产量提高 170%,乳酸含量降低
三、共培养中的种群分析方法
群体分析对于理解相互作用机制、环境响应及细胞功能具有重要意义。
3.1 分析方法概述
目前存在多种分析方法,从传统的细胞计数到先进的组学技术,可分为以下几类:
表1.微生物共培养分析的主要方法分类及应用
| 方法类别 | 具体技术 | 主要应用 |
|---|---|---|
| 细胞水平 | 平板计数、流式细胞术 | 分析细胞生理状态和群体动态 |
| 组学技术 | 宏基因组学、转录组学、蛋白质组学、代谢组学 | 揭示微生物间的相互作用信号 |
| 成像技术 | 显微图像分析、荧光显微镜 | 整合形态、荧光等多信号分离细胞 |
注:宏基因组学、转录组学和蛋白质组学的联合应用(如代谢组学、RNA测序以及荧光标记蛋白)能够高效地深入揭示微生物间的相互作用信号。
3.2 显微图像分析
3.2.1 基本原理
显微图像分析能够整合多种技术(荧光、吸光度、形态学、尺寸等),通过不同信号对细胞进行分离。
3.2.2 应用方式
表2.不同显微成像技术的适用条件与示例
| 方式 | 适用条件 | 示例 |
|---|---|---|
| 相差显微镜 | 微生物形态差异明显 | 监测共培养过程 |
| 荧光显微镜 | 基因工程标记或荧光染色 | 确定共培养组成成分 |
3.2.3 自动化与智能化
挑战:手动分析大量显微图像工作量巨大
当前解决方案:
机器学习算法:开发自动化细胞成像技术,实现数据自动解读
深度学习(神经网络):处理海量图像,是目前细胞检测领域的最先进方法
注意:当细胞的大小和形状存在差异,或图像背景变化较大时,自动化方法仍面临挑战,需要人工对图像特征进行定义。
3.3 组学分析
3.3.1 宏基因组学
作用:
深入揭示微生物的生理状态及其与环境之间的相互作用
理解微生物如何摄取和降解营养物质
解析微生物在面对压力时如何调整代谢途径
3.3.2 代谢组学
作用:追踪表型和细胞功能的动态变化
主要挑战:
基因与代谢物之间存在巨大鸿沟,制约了知识获取深度
难以全面分析细胞内和细胞外的所有代谢物(代谢物化学结构和性质差异极大)
代谢物浓度和组成会迅速响应环境刺激,离线分析可能导致信息丢失
应对策略:多组学联合分析可提供更深层次的洞见
3.3.3 转录组分析
常用技术:实时定量PCR(qPCR)
应用:检测和定量生物量及目标DNA序列
优势:与其他组学方法结合,结果可与种群分析相关联
3.4 各类共培养的种群监测方法
3.4.1 酵母共培养
表3. 酵母共培养体系中用于监测菌群数量的分析方法
| 分析方法 | 物种 |
|---|---|
| 平板计数法(选择性) | Torulaspora delbrueckii Zymaflore™ alpha 与 Saccharomyces cerevisiae QA23 |
| 平板计数法(选择性) | Torulaspora delbrueckii Biodiva 与 Saccharomyces cerevisiae Safale S-04 |
| 平板计数法(非选择性) | Wickerhamomyces anomalus Y3604 与 Saccharomyces cerevisiae Y3401 |
| 平板计数法(选择性) | Cyberlindnera fabianii 65 与 Saccharomyces cerevisiae Nottingham™ |
| 代谢物分析(HS-SPME–GC-MS) | Lallemand(注:此处指由 Lallemand 公司提供的酵母菌株或混合培养体系;原文未明确具体物种) |
| 代谢物分析(HPLC) | Aureobasidium pullulans CCY 27-1-94 与 Saccharomyces cerevisiae 11,982 |
| 代谢物分析(HPLC) | Candida tropicalis TISTR 5306 与 Saccharomyces cerevisiae TISTR 5606 |
| 代谢物分析(HPLC) | Scheffersomyces stipitis 与 Saccharomyces cerevisiae PE-2 |
3.4.2 酵母-细菌共培养
表4. 酵母–细菌共培养体系中用于监测菌群数量的分析方法
| 分析方法 | 物种 (Species) |
|---|---|
| 平板计数(非选择性) | 酿酒酵母 (Saccharomyces cerevisiae) NTUAFM-Y027 和 苹果汉逊酵母 (Hanseniaspora uvarum) NTUAFM-Y011 |
| 平板计数(选择性和非选择性) | 乳酸片球菌 (Pediococcus acidilactici) UFLA BFFCX 27.1,嗜酸乳杆菌 (Lactobacillus acidophilus) LACA 4,和 酿酒酵母 (Saccharomyces cerevisiae) UFLA YFFBM 18.03 |
| qPCR 平板计数(选择性) 代谢物分析(重量分析和 UPLC) | 鼠李糖乳杆菌 (Lactobacillus rhamnosus) ATCC 9595, R0011, 和 RW-9595 M 与 酿酒酵母 (Saccharomyces cerevisiae) Instaferm®, Lallemand |
| 平板计数(选择性) 代谢物分析(HS-SPME-GC-MS) | 植物乳植杆菌 (Lactiplantibacillus plantarum) L6 和 粘红酵母 (Rhodotorula mucilaginosa) Y9 |
| 平板计数(选择性) 代谢物分析(HPLC) | 副干酪乳杆菌 (Lactobacillus paracasei) L26 和 酿酒酵母 (Saccharomyces cerevisiae) S-04 |
| 平板计数(选择性) 代谢物分析(HPLC) | 酿酒酵母 (Saccharomyces cerevisiae) 和 巴氏乳杆菌 (Lactobacillus panis) |
| 平板计数(选择性) 代谢物分析(LC-MS) | 汉逊德巴利酵母 (Debaryomyces hansenii) D1,木糖葡萄球菌 (Staphylococcus xylosus) S1,和 戊糖片球菌 (Pediococcus pentosaceus) PP1 |
| 平板计数(非选择性) | 植酸梭菌 (Clostridium phytofermentans) ISDgT,摩氏假丝酵母 (Candida molischiana) Y-2237,和 酿酒酵母 (Saccharomyces cerevisiae) cdt-1 |
| 平板计数(非选择性) 代谢物分析(GC-MS 和 HPLC) | 大肠杆菌 (Escherichia coli) MG1655 和 酿酒酵母 (Saccharomyces cerevisiae) BY4700 |
| 平板计数(选择性) 代谢物分析(琼脂扩散试验和 GC-MS) 代谢物分析(HPLC) | 乳酸乳球菌 (Lactococcus lactis) ATCC 11,454 和 酿酒酵母 (Saccharomyces cerevisiae) W303-1 A |
| 代谢物分析(UPLC-MS) | 大肠杆菌 (Escherichia coli) BL21(DE3) 和 酿酒酵母 (Saccharomyces cerevisiae) BY4741 |
| qPCR 代谢物分析(HS-GC-HTIMS) | 大肠杆菌 (Escherichia coli) AN2014 和 毕赤酵母 (Komagataella phaffii) (Pichia pastoris B52) |
| qPCR 二代测序 代谢物分析(HS-GC-IMS) | 克氏乳杆菌 (Lactobacillus kefiranofaciens),开菲尔乳杆菌 (Lactobacillus kefiri),肠膜明串珠菌 (Leuconostoc mesenteroides),乳酸乳球菌 (Lactococcus Lactis),马克斯克鲁维酵母 (Kluyveromyces marxianus),苏黎世卡萨克酵母 (Kazachstania turicensis),单孢卡萨克酵母 (Kazachstania unispora),和 异常德克酵母 (Dekkera anomalous) |
| 平板计数(选择性和非选择性) 宏基因组鸟枪法测序 代谢物分析(HPAEC-PAD、HPLC-MSMS 和 HS/SPME-GC-MS) | 动物双歧杆菌 (Bifidobacterium animalis),乳杆菌属 (Lactobacillus sp.),乳球菌属 (Lactococcus sp.),慢乳杆菌属 (Lentilactobacillus sp.),明串珠菌属 (Leuconostoc sp.),链球菌属 (Streptococcus sp.),异常德克酵母 (Dekkera anomola),苏黎世卡萨克酵母 (Kazachstania turicensis),单孢卡萨克酵母 (Kazachstania unispora),马克斯克鲁维酵母 (Kluveromyces marxianus),和 膜醭毕赤酵母 (Pichia membranifaciens) |
| qPCR 代谢物分析(RF-HPLC) 细胞大小分布(库尔特计数器) 代谢物分析(HPLC) 流式细胞术 荧光报告基因 | 运动发酵单胞菌 (Zymomonas mobilis),水双歧杆菌 (Bifidobacterium aquikefiri),大麦酒乳杆菌 (Liquorilactobacillus hordei),萨氏酒乳杆菌 (Liquorilactobacillus satsumensis),纳氏酒乳杆菌 (Liquorilactobacillus nagelii),苏尼奥明串珠菌 (Leuconostoc suionicum),布鲁塞尔酒香酵母 (Brettanomyces bruxellensis),酿酒酵母 (Saccharomyces cerevisiae),真巴氏酵母 (Saccharomyces eubayanus),和 德尔布氏酵母 (Torulaspora delbrueckii) 等 |
| 动物双歧杆菌 (Bifidobacterium animalis) BB04 和 异常威克汉姆酵母 (Wickerhamomyces anomalus) Y-5 | |
| 瓦氏汉逊酵母 (Hanseniaspora valbyensis) Hs-0302 和 肠膜明串珠菌 (Leuconostoc mesenteroides) Le-0304 | |
| 酿酒酵母 (Saccharomyces cerevisiae) BY4741 和 枯草芽孢杆菌 (Bacillus subtilis) W168 |
3.4.3 细菌共培养
表5.细菌共培养体系中用于监测菌群数量的分析方法
| 分析方法 | 物种 |
|---|---|
| 平板计数(选择性和非选择性) | Methyloparacoccus murrellii LMG 27,482 和 Cupriavidus necator LMG 1201 (默氏甲基对球菌 LMG 27,482 和 食酸罗尔斯通菌 LMG 1201) |
| 平板计数(选择性和非选择性) | Lactobacillus kefiranofaciens HL1 和 Lactococcus lactis APL15 (克氏乳杆菌 HL1 和 乳酸乳球菌 APL15) |
| 平板计数(选择性和非选择性) | Pseudomonas protegens JZ R-192 与 Listeria monocytogenes DSM 2060 以及 Acinetobacter johnsonii JZ RK-116 与 Carnobacterium maltaromaticum JZ RK-71 (保护性假单胞菌 JZ R-192 与 单核细胞增生李斯特菌 DSM 2060 以及 约翰逊不动杆菌 JZ RK-116 与 麦芽香肉杆菌 JZ RK-71) |
| 平板计数(非选择性) | Propionibacterium freudenreichii NCC 1177 和 Bacillus amyloliquefaciens NCC 156 (弗雷德里希丙酸杆菌 NCC 1177 和 解淀粉芽孢杆菌 NCC 156) |
| 代谢物分析(HPLC) | |
| 平板计数(选择性和非选择性) | Lactococcus lactis FM03-V1 和 Leuconostoc mesenteroides FM06 (乳酸乳球菌 FM03-V1 和 肠膜明串珠菌 FM06) |
| qPCR | |
| qPCR | |
| 代谢物分析(HPLC 和 HS-SPME GC-MS) | |
| 平板计数(选择性和非选择性) | Staphylococcus aureus ATCC 29,213 和 Enterococcus faecalis 41FL1 (金黄色葡萄球菌 ATCC 29,213 和 粪肠球菌 41FL1) |
| 代谢物分析(酶免疫测定) | |
| qPCR | |
| qPCR | Clostridium cellulovorans 743B 和 Clostridium beijerinckii NCIMB 8052 (嗜纤维梭菌 743B 和 拜氏梭菌 NCIMB 8052) |
| 代谢物分析(琼脂扩散试验和 HPLC) | Komagataeibacter xylinum ATCC 700,178 和 Lactococcus lactis ATCC 11,454 (木醋杆菌 ATCC 700,178 和 乳酸乳球菌 ATCC 11,454) |
| qPCR | |
| 代谢物分析(HPLC) | Gluconobacter oxydans 和 Ketogulonicigenium vulgare (氧化葡萄糖酸杆菌 和 普通酮古龙酸菌) |
| qPCR | |
| 平板计数(选择性) | Bifidobacterium breve UCC2003 和 Bifidobacterium bifidum PRL2010 (短双歧杆菌 UCC2003 和 两歧双歧杆菌 PRL2010) |
| 代谢物分析(HPAEC-PAD) | |
| DNA 微阵列分析 | |
| 代谢物分析(13C 同位素标记和 LC-MS/MS) | Lactobacillus delbrueckii subsp. bulgaricus ATCC BAA-365, 和 Streptococcus thermophilus LMG 18,311 (德氏乳杆菌保加利亚亚种 ATCC BAA-365, 和 嗜热链球菌 LMG 18,311) |
| 代谢物分析(HPTLC) | Escherichia coli DSM 18,039, Bacillus subtilis 29,784, Bacillus subtilis MIC39, 和 Bacillus subtilis ATCC 6633 (大肠杆菌 DSM 18,039, 枯草芽孢杆菌 29,784, 枯草芽孢杆菌 MIC39, 和 枯草芽孢杆菌 ATCC 6633) |
| 代谢物分析(HPLC) | Megasphaera elsdenii ATCC 25,940 和 Lactobacillus pentosus ATCC 8041 (埃氏巨球型菌 ATCC 25,940 和 戊糖乳杆菌 ATCC 8041) |
| 代谢物分析(HPLC) | Kosakonia oryzendophytica FY-07 和 Leuconostoc carnorum 4010 (稻内生科萨克氏菌 FY-07 和 肉明串珠菌 4010) |
| 代谢物分析(UHPLC-MS/MS) | Serratia plymuthica 4Rx13, AS9 和 Bacillus subtilis B2g (普城沙雷氏菌 4Rx13, AS9 和 枯草芽孢杆菌 B2g) |
| 代谢物分析(气体流速) | Clostridium carboxidivorans DSM 15,243 和 Clostridium kluyveri DSM 555 (嗜碳梭菌 DSM 15,243 和 克氏梭菌 DSM 555) |
| FISH-流式细胞术 | |
| 代谢物分析(HPLC) | Escherichia coli W3110 和 Corynebacterium glutamicum DM1729 (大肠杆菌 W3110 和 谷氨酸棒杆菌 DM1729) |
| 荧光显微镜 | |
| 流式细胞术 | Geobacter sulfurreducens PCA (DSMZ 12127) 和 Shewanella oneidensis MR1 (ATCC 700550) (还原地杆菌 PCA (DSMZ 12127) 和 奥奈氏希瓦氏菌 MR1 (ATCC 700550)) |
| 共聚焦激光扫描显微镜 | |
| 流通池共聚焦激光扫描显微镜 | Streptococcus mutans UA159 serotype c (ATCC 700610), Streptococcus oralis J22, 和 Candida albicans SN250 (变形链球菌 UA159 c血清型 (ATCC 700610), 口腔链球菌 J22, 和 白色念珠菌 SN250) |
3.5 基于代谢物的种群分析
3.5.1 通用特点
许多研究分析了糖类、有机酸和乙醇,揭示了部分代谢通量。然而,往往难以将合成过程归因于特定的微生物种类。
3.5.2 特异性代谢标志物
表6.共培养体系中关键代谢标志物及其指示意义
| 标志物 | 产生菌 | 应用价值 |
|---|---|---|
| 胞外多糖 | 乳酸菌 | 分泌到胞外,含量可反映代谢状况 |
| Nisin(尼辛) | 乳酸乳球菌 | 独特的抗菌肽,浓度可评估代谢活性 |
| 酚类物质 | 鼠李糖乳杆菌 | 应激条件下利用酚酸作为电子受体,指示代谢活跃程度 |
| 氨基酸 | 多种菌 | 特定氨基酸消耗可反映菌体生长状态 |
3.5.3 代谢物分析与基因表达联用
多项研究采用 qPCR 结合代谢物分析 的方法,探究与代谢物转化和合成密切相关的基因。
典型案例:
测定紫杉烷类化合物(大肠杆菌产生)
测定含氧紫杉烷类化合物(酿酒酵母产生)
测定乙酸(大肠杆菌产生、酿酒酵母消耗)
局限性:紫杉二烯和含氧紫杉烷的生物合成过程复杂,限制了代谢物分析评估种群生长的能力。
3.6 流式细胞术在种群分析中的应用
由于细胞大小存在差异,流式细胞术分析成为可能。
典型研究:
在酿酒酵母 BY4741 与枯草芽孢杆菌 W168 之间建立受控的信号传递
将能够检测酿酒酵母产生的群体感应肽的荧光报告基因整合到枯草芽孢杆菌中
利用流式细胞术检测肽类物质,实现单细胞水平的相互作用分析
3.7 特殊应用:生物膜群体分析
3.7.1 地杆菌与希瓦氏菌共培养
研究目标:探究生物电化学系统中生物膜的形成过程
方法:
共聚焦激光扫描显微镜分析生物膜
荧光信号测定生物膜平均厚度
流式细胞术分别计数浮游细胞和生物膜中的细胞
主要发现:
共培养形成的生物膜比地杆菌单菌培养的生物膜厚 35%(后者几乎完全由地杆菌组成)
浮游状态的希瓦氏菌细胞对地杆菌产生积极影响
推测机制:希瓦氏菌释放的 H₂ 和黄素作为额外能量来源,促进电子转移效率;裂解释放的胞外 DNA 有助于生物膜形成与稳定
3.7.2 变异链球菌与念珠菌共培养
技术:
共聚焦激光显微镜 + 图像处理技术
流动池微流控装置
成果:
实现生物膜中群体生长的实时监测
捕捉单个细胞的演化过程
将细胞集群和聚集体归类为结构化的群落
3.8 综合分析与展望
借助种群分析对微生物功能及其相互作用的深入研究,我们可以:
✅ 充分挖掘微生物的代谢潜力
✅ 开发新型生物活性代谢物
✅ 精准调控代谢流,实现特定终产物的高效生产
然而,上述方法的结合虽然有助于更深入地理解群体,但所提供的信息仍然有限。亟需开发适用于在线和准在线的单细胞监测方法,以实现对微生物生长的有效控制。
四、单细胞监测
单细胞监测是实现在线/近线控制生长的先决条件。通过单细胞水平的分析,我们能够实时掌握微生物的生长状态、形态变化和生理活动,从而实现对培养过程的精准调控(见图2)。
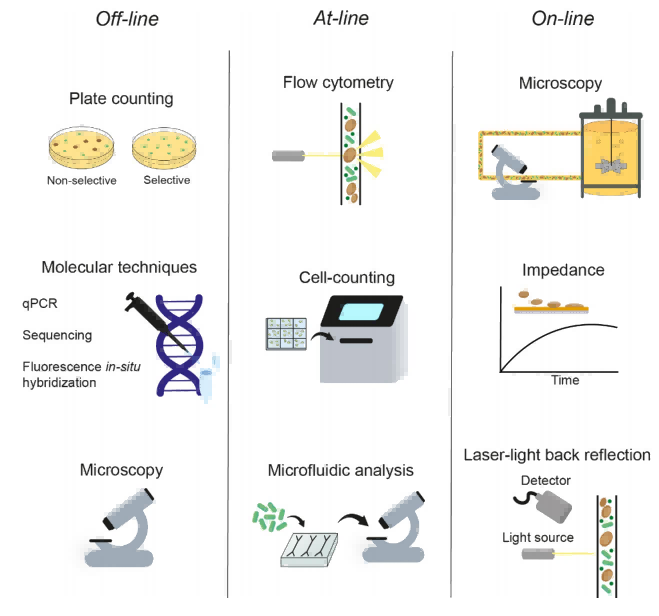
图2. 离线、在线及在线单细胞监测。离线方法包括平板计数、分子技术和显微镜观察。在线旁线方法包括流式细胞术、自动细胞计数以及使用流动室的显微镜观察。最后,在线方法包括连接生物反应器的显微镜、阻抗法和激光背向反射。
4.1 主要技术
表7.单细胞监测的主要技术特点与应用案例
| 技术 | 特点 | 应用案例 |
|---|---|---|
| 自动化图像分析 | 监测生长、评估形态、选择性计数 | 酵母、乳酸菌无菌培养 |
| 原位显微镜 | 测定细胞密度和活性,无需取样 | 较传统方法更具优势 |
| 在线流式细胞术 | 长期、高频测量,不遗漏瞬时事件 | 乳酸嗜酸菌发酵 |
| 荧光原位杂交 | 特异性 23S rRNA 探针标记 | 精确测定细胞数量 |
| 软传感器 | 偏最小二乘法关联 OD 扫描谱与生物量 | 共培养组成测定 |
| 微流控平台 | 连续成像、实时评估 | 铜绿假单胞菌+白色念珠菌互作 |
4.2 典型案例
案例1:散射光谱在线监测
Geinitz 等采用 48 孔板测量 280~720 nm 散射光谱,结合偏最小二乘回归估算克鲁维酵母与乳酸乳球菌共培养的群体组成,偏差约 25%。
案例2:葡萄糖脉冲策略稳定种群
Martinez 等利用葡萄糖脉冲策略调节酿酒酵母与大肠杆菌共培养的种群组成,通过在线流式细胞术验证,模型预测与实验结果吻合。
4.3分析方法汇总
表8.用于共培养群体分析的分析方法总结(平板计数、细胞大小分布、代谢物分析、转录组学、显微镜检查和流式细胞术)
| 分析方法 | 优点 | 局限性 |
|---|---|---|
| 平板计数 | 检测到活细胞 | 分析时间长 无法检测到休眠细胞、对氧敏感的细胞和病原体 提供的关于微生物类型的信息有限 结果可能取决于培养基成分 |
| 细胞大小分布(库尔特计数器) | 快速 重复性好 | 如果细胞大小重叠,可能会高估和/或低估细胞数量 |
| 代谢物分析(特别是色谱法) | 稳健 可靠 可转移的分析方法 | 代谢物的丰度取决于培养条件 只能分析稳定、可提取、可分离和可检测的代谢物 如果使用离线方法,分析时间会延长 可能需要从培养基中分离和化学转化 |
| 转录组学(主要是qPCR 和测序) | 灵敏 靶向特异性 单细胞分析 | 无法区分死细胞和活细胞 仅能检测已知序列 |
| 流式细胞术 | 单细胞分析 亚群分析 最终可能适用于在线选项 | 如果使用荧光染色,既耗时又容易出错 当使用多种荧光团来区分微生物时,存在发射光谱物理重叠的风险 |
| 显微镜检查 | 单细胞分析 接近实时分析(如果是在线/原位显微镜) 基于图像的方法,识别可以自动化 | 仅适用于具有独特形态的微生物 如果使用荧光染色,既耗时又容易出错 |
五、结论与展望
5.1 核心结论
微生物共培养研究具有重要现实意义,可带来以下优势:
提高生产效率
增强菌群稳定性
探索鲜为人知的代谢产物合成途径
当前瓶颈:对共培养过程可控性的认知尚显不足,制约了工业化应用。
5.2 实现工业化应用的价值
表9.共培养体系的主要收益维度与具体优势
| 收益维度 | 具体表现 |
|---|---|
| 底物适应性 | 可利用更多种类的原料,包括废弃物 |
| 产品收率 | 优化菌群比例,提高产量 |
| 产品品质 | 风味、口感、营养价值更稳定 |
| 地域扩展性 | 在基础设施薄弱地区也能推广应用 |
| 废弃物增值 | 将生物源废弃物转化为高附加值产品 |
5.3 最终愿景
✅ 传统共培养工艺可实现工业化转移
✅ 转移过程不降低产品质量
✅ 满足日益增长的健康食品需求
✅ 支持区域循环经济,推动可持续产品发展
参考文献
1.Heins A, Hoang MD, Weuster-Botz D (2022) Advances in automated real‐time flow cytometry for monitoring of bioreactor processes. Eng Life Sci 22(3–4):260–278
2.Jia X, Xu J, Liu L, Liang X, He Y, Han L et al (2024) Strategies for enhancing short-and medium-chain fatty acids synthesis from corn stover-based sugar: A new perspective on co-culture. J Environ Chem Eng 12(6):114836
3.Kruse S, Becker S, Pierre F, Morlock GE (2023) Metabolic profiling of bacterial co-cultures reveals intermicrobiome interactions and dominant species. J Chromatogr A 1694:463911
4.Latif A, Shehzad A, Niazi S, Zahid A, Ashraf W, Iqbal MW et al (2023) Probiotics: mechanism of action, health benefits and their application in food industries. Front Microbiol 14:1216674
5.Liu Y, Nie R, Shen K, Diao X, Liu G (2024) Multi-omics profiling reveals the molecular mechanism of Bifidobacterium animalis BB04 in co-culture with Wickerhamomyces anomalus Y-5 to induce bifidocin A synthesis. World J Microbiol Biotechnol 40(11):1–17
相关产品
HZB366353:鼠李糖乳杆菌 | Lacticaseibacillus rhamnosus
HZB231920:酿酒酵母 CEN.PK2-1D | Saccharomyces cerevisiae CEN.PK2-1D
HZB224707:动物双歧杆菌乳亚种 | Bifidobacterium animalis subsp. lactis
HZB218425:运动发酵单胞菌 | Zymomonas mobilis
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-05-22
编制人:思琪
审稿人:叶凡