食源性细菌病原体在检测和预防方面的新兴方法
来源:武汉市灰藻生物科技有限公司 浏览量:136 发布时间:2026-04-22 13:17:12
一、现状与挑战:严峻的公共卫生议题
1.1 疾病负担与经济损失
食源性细菌病原体依然是全球公共卫生的重大威胁。据世界卫生组织(WHO)估计:
患病人数:每年约有 6亿人 患食源性疾病。
死亡人数:导致约 42万人 死亡,其中五岁以下儿童占死亡人数的近 30%。
经济影响:包括治疗费用、生产力损失及贸易中断,仅美国每年成本就超150亿美元。
1.2 主要致病菌及其威胁
表1.主要食源性细菌病原体及其健康风险与特征
| 病原体 | 主要关联食品 | 健康风险与特征 |
|---|---|---|
| 沙门氏菌 (Salmonella) | 家禽、蛋类、乳制品 | 胃肠炎主要病因,具有高流行率。 |
| 大肠杆菌 O157:H7 | 未煮熟牛肉、生奶 | 产生志贺毒素,可导致溶血性尿毒症综合征 (HUS)。 |
| 单增李斯特菌 (L. monocytogenes) | 即食食品 ( RTE ) | 冷藏条件下仍可生长,孕妇、新生儿死亡率高。 |
| 弯曲杆菌 (Campylobacter) | 未煮熟家禽 | 微需氧菌,与吉兰-巴雷综合征相关。 |
| 肉毒梭菌 (C. botulinum) | 罐头、发酵食品 | 产生神经毒素,导致肌肉麻痹甚至死亡。 |
1.3 传统方法的局限性
传统的基于培养、生化和免疫学的检测方法虽然可靠,但存在明显短板:
耗时长:通常需要2-7天。
漏检风险:难以检测"活的但不可培养" (VBNC) 状态的病原体。
适应性差:病原体正迅速适应环境压力(如耐冷、耐酸),且多重耐药菌株呈上升趋势。
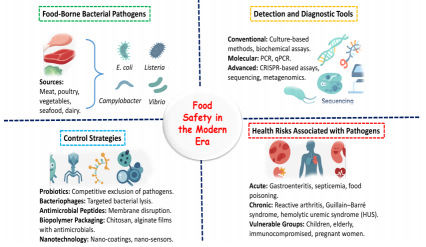
图1.食源性致病菌的综合管理:从源头到健康影响
二、微生物学机制:病原体如何生存与致病
2.1 分类学、生理学和毒力因子
○ 沙门氏菌 (Salmonella)
分类学:肠杆菌科,革兰氏阴性杆菌,包括超过2600个血清型
生理学:兼性厌氧菌,最适生长温度37℃,pH范围4.0-9.0,耐胆汁盐
毒力因子:
SPI-1:编码III型分泌系统,介导上皮细胞入侵
SPI-2:促进在巨噬细胞内的存活和复制
鞭毛:提供运动性,参与生物膜形成
脂多糖(LPS):内毒素,引发强烈免疫反应
○ 大肠杆菌 O157:H7
分类学:肠杆菌科,肠杆菌属,致病性大肠杆菌(EHEC)亚群
生理学:兼性厌氧,37℃最适温度,能耐受pH 2.0-8.0,在低温下仍可缓慢生长
毒力因子:
志贺毒素(Stx):抑制蛋白质合成,导致细胞死亡
LEE毒力岛:编码III型分泌系统和Intimin蛋白,形成"附着-消除"病变
溶血素:破坏红细胞,促进细菌扩散
菌毛:介导肠道定植
○ 单增李斯特菌 (Listeria monocytogenes)
分类学:李斯特菌科,革兰氏阳性短杆菌,兼性胞内寄生
生理学:兼性厌氧,能在0-45℃生长(嗜冷菌),耐盐、耐酸,在pH 4.4-9.6可存活
毒力因子:
李斯特溶血素O(LLO):溶细胞素,破坏吞噬体膜
ActA蛋白:促进肌动蛋白聚合,实现细胞间传播
磷脂酶:帮助细菌在细胞内逃逸
InlA/InlB蛋白:介导上皮细胞和肝细胞入侵
○ 弯曲杆菌 (Campylobacter)
分类学:弯曲杆菌科,革兰氏阴性螺旋形杆菌
生理学:微需氧菌(5-10% O₂),最适生长温度42℃(禽类体温),对氧敏感
毒力因子:
鞭毛:提供螺旋运动能力,是主要的毒力因子
细胞致死性膨胀毒素(CDT):诱导DNA损伤和细胞周期阻滞
外膜蛋白:介导黏附和侵入
脂寡糖(LOS):类似LPS,引发炎症反应
○ 肉毒梭菌 (Clostridium botulinum)
分类学:梭菌科,革兰氏阳性厌氧杆菌,能形成芽孢
生理学:严格厌氧,最适温度30-37℃,产生强效神经毒素
毒力因子:
肉毒毒素:已知最毒的生物毒素,阻断神经肌肉接头
溶血素:破坏红细胞和组织
蛋白酶:帮助毒素释放和激活
2.2 毒力因子的调控机制
温度调控:沙门氏菌的SPI-1在37℃表达,25℃时表达鞭毛基因
pH调控:大肠杆菌O157:H7在酸性环境下上调酸耐受反应基因
群体感应:通过AI-2信号分子协调生物膜形成和毒力表达
双组分系统:如李斯特菌的VirRS系统调控LLO表达
2.3 环境生存策略
生物膜形成:在食品加工设备表面形成胞外聚合物 (EPS),对抗生素和消毒剂的抵抗力显著增强。
应激反应:
酸耐受:大肠杆菌和沙门氏菌穿越胃酸屏障的关键。
冷适应:单增李斯特菌通过PrfA调控子在冷藏食品中持续存在。
芽孢:肉毒梭菌和产气荚膜梭菌的芽孢对高温、干燥有极强抗性。
2.4 适应性机制的临床意义
表2不仅突出了这些病原体的临床意义,还展示了它们通过生物膜形成、孢子产生及耐受应激等机制实现适应性的能力,这使得检测与控制策略更加复杂。这些适应性机制与图2中概述的感染途径和毒力网络相互印证,共同构成了食源性病原体威胁公众健康的复杂图景。
表2.主要食源性细菌病原体:分类学、疾病、毒力因子及生存策略
| 病原体 | 分类学 (属, 科) | 常见食物来源 | 主要疾病 (急/慢性) | 毒力因子 | 生存策略 (生物膜, 芽孢, 应激适应) | 易感人群 |
|---|---|---|---|---|---|---|
| 肠炎沙门氏菌 (Salmonella enterica) | 肠杆菌科 | 禽肉, 鸡蛋, 农产品 | 沙门氏菌病 (胃肠炎) | III型分泌系统 (T3SS), 内毒素, 粘附蛋白 | 酸耐受, 表面生物膜 | 儿童, 老年人 |
| 大肠杆菌 O157:H7 (Escherichia coli O157:H7) | 肠杆菌科 | 牛肉, 蔬菜, 牛奶 | 出血性结肠炎, 溶血性尿毒综合征 (HUS) | 志贺毒素, 菌毛, 粘附素 | 酸耐受, 活的非可培养 (VBNC) 状态 | 儿童 |
| 单核细胞增生李斯特菌 (Listeria monocytogenes) | 李斯特菌科 | 乳制品, 即食肉类 | 李斯特菌病 (脑膜炎) | 李斯特溶血素O (LLO), 内化素 | 冷耐受, 生物膜 | 孕妇, 新生儿 |
| 空肠弯曲杆菌 (Campylobacter jejuni) | 弯曲杆菌科 | 禽肉, 牛奶 | 弯曲杆菌病, 吉兰-巴雷综合征 | CDT毒素, 粘附素, 运动性 | 微需氧, VBNC | 普通人群, 免疫受损者 |
| 肉毒梭菌 (Clostridium botulinum) | 梭菌科 | 罐头食品, 鱼类 | 肉毒中毒 | 神经毒素 (BoNTs) | 芽孢形成 | 所有人群, 尤其是老年人 |
| 坂崎克罗诺杆菌 (Cronobacter sakazakii) | 肠杆菌科 | 婴儿配方奶粉 | 新生儿脑膜炎, 败血症 | 荚膜, 铁载体 | 脱水耐受, 生物膜 | 新生儿, 婴儿 |
| 鲍氏不动杆菌 (Acrobacter butzleri) | 弧菌科 (注:原文可能有误,通常为不动杆菌属) | 禽肉, 乳制品 | 肠炎, 腹泻 | 粘附素, 毒素, 运动性 | 生物膜形成, 微需氧 | 普通人群 |
| 痢疾志贺氏菌 (Shigella dysenteriae) | 肠杆菌科 | 受污染的水, 蔬菜 | 痢疾, HUS | 志贺毒素, 侵袭性质粒, 抗原 | 应激适应, VBNC | 营养不良的儿童 |
| 霍乱弧菌 (Vibrio cholerae) | 弧菌科 | 受污染的水, 海鲜 | 霍乱 (急性水样腹泻) | 霍乱毒素, TCP菌毛 | VBNC, 生物膜 | 儿童, 卫生条件差地区 |
| 副溶血性弧菌 (Vibrio parahaemolyticus) | 弧菌科 | 生海鲜, 贝类 | 急性胃肠炎 | TDH, TRH毒素, 溶血素 | 盐耐受, VBNC | 海鲜消费者 |
| 嗜水气单胞菌 (Aeromonas hydrophila) | 气单胞菌科 | 海鲜, 蔬菜 | 胃肠炎, 伤口感染 | 溶血素, 肠毒素 | 生物膜形成, 应激适应 | 免疫受损个体 |
| 肠结肠耶尔森菌 (Yersinia enterocolitica) | 肠杆菌科 | 猪肉, 牛奶, 未经处理的水 | 耶尔森菌病 (肠系膜淋巴结炎) | 侵袭素, YadA粘附素 | 冷耐受, 生物膜 | 儿童, 老年人 |
| 蜡样芽孢杆菌 (Bacillus cereus) | 芽孢杆菌科 | 米, 意面, 乳制品 | 食物中毒 (呕吐, 腹泻) | 肠毒素, 蜡样杆菌毒素 | 芽孢形成, 应激适应 | 所有年龄段 |
| 金黄色葡萄球菌 (Staphylococcus aureus) | 葡萄球菌科 | 乳制品, 肉类, 沙拉 | 葡萄球菌性食物中毒 | 肠毒素, 超抗原 | 生物膜, 毒素稳定性 | 所有年龄段 |
| 产气荚膜梭菌 (Clostridium perfringens) | 梭菌科 | 煮熟的肉, 炖菜 | 食物中毒 (腹泻) | 肠毒素 (CPE) | 芽孢形成 | 所有年龄段 |
| 粪肠球菌 (Enterococcus faecalis) | 肠球菌科 | 奶酪, 发酵食品 | 机会性感染, 菌血症 | 聚集物质, 细胞溶解素 | 生物膜, 抗生素耐药性 | 免疫受损者 |
| 幽门螺杆菌 (Helicobacter pylori) | 螺杆菌科 (Helicobacteraceae) | 牛奶,受污染的水 | 胃炎,胃癌 | CagA, VacA 毒素 | 酸适应,生物膜 | 成人,高危人群 |
| 铜绿假单胞菌 (Pseudomonas aeruginosa) | 假单胞菌科 (Pseudomonadaceae) | 肉类,乳制品,医院食品 | 机会性感染,败血症 | 弹性蛋白酶,外毒素 A | 生物膜,抗生素耐药性 | 住院患者 |
| 迟缓爱德华氏菌 (Edwardsiella tarda) | 肠杆菌科 (Enterobacteriaceae) | 鱼类,水生食品 | 胃肠炎,败血症 | 溶血素,铁载体 | 生物膜,VBNC (活的非可培养状态) | 普通人群 |
| 布鲁氏菌属 (Brucella spp.) | 布鲁氏菌科 (Brucellaceae) | 未经巴氏消毒的乳制品,肉类 | 布鲁氏菌病(全身性感染) | 脂多糖,脲酶 | 细胞内生存 | 农村和农场人群 |
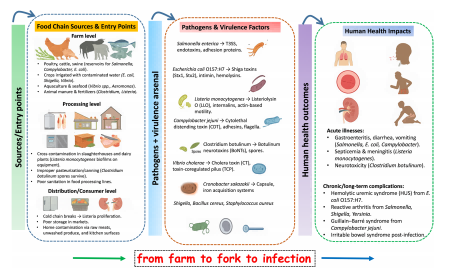
图2.食源性细菌病原体的感染途径
注:重点展示了其来源、毒力因子,以及从急性胃肠炎到慢性并发症的相关健康影响,并涵盖了抗菌药物耐药性和气候变化等全球性驱动因素。
三、检测技术革新:从分子诊断到智能预测
传统的微生物学检测方法依赖于培养基的增菌、分离与生化鉴定,虽然作为金标准能够分离活性菌株,但通常耗时2-7天,对低丰度或活而不可培养(VBNC)状态的病原体敏感性不足,且易受竞争性微生物干扰。为突破这些局限,现代检测技术正朝着快速、精准、智能化的方向加速演进。
3.1 快速分子检测技术
○ PCR/qPCR:目前的常用方法,如针对沙门氏菌的invA基因、单增李斯特菌的hlyA基因检测。qPCR具备定量能力,经富集后可检测每克食品中低至10-100 CFU的大肠杆菌O157:H7,已成为监管检测的核心工具。
○ 区分活细胞技术:
● PMA/EMA染料:结合PCR,选择性抑制膜受损死细胞的DNA扩增,富集活菌信号。
● RT-PCR:以短寿命mRNA为靶标,反映细胞代谢活性,精准识别即食食品中具有活性的单增李斯特菌等病原体。
○ CRISPR诊断:SHERLOCK和DETECTR平台利用Cas酶的序列特异性切割能力,实现飞摩尔级超灵敏检测。与RPA/LAMP等恒温扩增技术联用后,可在1小时内完成牛奶中沙门氏菌的现场检测,具备高特异性和便携性。
3.2 基因组学与宏基因组学
● 全基因组测序(WGS):作为疫情溯源的金标准,能够全面解析病原体基因组,精准追踪污染源。例如,在冷冻蔬菜中单增李斯特菌疫情和家禽沙门氏菌暴发事件中,WGS成功实现了菌株水平的溯源分析。Illumina、牛津纳米孔等平台还支持质粒与可移动遗传元件的分析,助力耐药基因传播研究。
● 宏基因组学:采用shotgun测序策略,无需培养即可揭示复杂食品基质中的完整微生物群落结构,有效识别低丰度或VBNC状态的病原体,如家禽加工环境中的弯曲菌。尽管存在成本高、难以区分活死细胞等挑战,但随着计算分析流程的优化,其实用性正不断提升。
3.3 生物信息学与人工智能(AI)
● 数据库与网络:GenBank、ENA等公共数据库与GenomeTrakr、PulseNet等专业系统,实现了全球范围内的基因组数据共享与比对,为跨国疫情追踪提供支撑。
● 机器学习与预测模型:
○ 利用随机森林、XGBoost等算法,整合基因组、环境与生产数据,预测病原体生长响应、耐药性及加工环境中的持续污染风险。
○ 预测性微生物学模型(如ComBase、PMP)结合AI技术,可模拟不同温度、pH和储存条件下的微生物行为,实现定量微生物风险评估(QMRA)。
○ 通过整合WGS监测与机器学习框架,已能近乎实时地识别暴发集群与污染热点,推动食品安全管理从被动响应向主动预警转型。
从传统培养到CRISPR快速检测,再到WGS溯源与AI智能预测,检测技术的迭代不仅大幅缩短了诊断周期,更构建起一个多维度、数据驱动的现代食品安全防控体系。表3和图3总结了这些方法的比较特征及时间演变历程。
表3.食源性病原体的传统、分子及先进检测方法
| 检测方法 | 原理 | 靶标病原体 | 灵敏度 | 检测时间 | 优点 | 缺点 |
|---|---|---|---|---|---|---|
| 基于培养(Culture - based) | 在选择性/鉴别培养基上生长 | 沙门氏菌,李斯特菌,大肠杆菌 | 中等 | 2 - 7 天 | 金标准,可获得分离株 | 缓慢,漏检 VBNC |
| 生化试验(API,VITEK) | 酶活性,糖发酵 | 肠杆菌科 | 中等 | 1 - 2 天 | 简单,标准化 | 假阳性/假阴性 |
| PCR/qPCR | 毒力基因的 DNA 扩增 | 沙门氏菌,李斯特菌,大肠杆菌 | 高(10 - 100 CFU) | 数小时 | 高灵敏度和特异性 | 易受基质抑制 |
| RT - PCR | RNA 转录本检测 | 活性沙门氏菌,李斯特菌 | 极高 | 数小时 | 检测活病原体 | 昂贵,复杂 |
| 多重 PCR | 多重引物用于多靶标扩增 | 沙门氏菌,大肠杆菌,李斯特菌 | 高 | 数小时 | 多病原体检测 | 交叉反应问题 |
| DNA 微阵列 | 探针杂交 | 多种细菌物种 | 高 | 1 - 2 天 | 大规模分析 | 昂贵,需要先验序列知识 |
| NGS(WGS) | 全基因组测序 | 所有食源性病原体 | 极高 | 1 - 2 天 | 菌株水平分辨率,疫情追踪 | 成本高,需要生物信息学分析 |
| 宏基因组学 | 所有 DNA 的鸟枪法测序 | 混合微生物群落 | 高 | 1 - 3 天 | 检测不可培养病原体 | 昂贵,分析复杂 |
| CRISPR - based 检测 | Cas12/13 旁切活性 | 沙门氏菌,大肠杆菌,李斯特菌 | 极高 | <1 小时 | 便携,实时 | 仍为新兴技术 |
| LAMP(环介导等温扩增) | 恒温下的 DNA 扩增 | 沙门氏菌,李斯特菌,弯曲杆菌 | 高(10 CFU) | 30 - 60 分钟 | 快速,不需要热循环仪 | 引物设计关键,非特异性扩增 |
| 免疫测定(ELISA,侧向流动) | 抗体 - 抗原结合 | 大肠杆菌,沙门氏菌,李斯特菌 | 中等到高 | 1 - 4 小时 | 廉价,广泛使用 | 可能需要富集,交叉反应 |
| 生物传感器(电化学,光学) | 病原体结合后的信号变化 | 沙门氏菌,李斯特菌,大肠杆菌,弧菌 | 高(单细胞水平可能) | 分钟至 2 小时 | 便携,灵敏 | 昂贵,需要设备校准 |
| MALDI - TOF 质谱 | 蛋白/肽质量指纹图谱 | 李斯特菌,沙门氏菌,大肠杆菌 | 高 | 分钟至数小时 | 快速,准确鉴定 | 依赖数据库,需要分离株 |
| 等温 RPA | 低温下的 DNA 扩增 | 大肠杆菌,沙门氏菌 | 高 | 20 - 40 分钟 | 低温恒温工作 | 对污染敏感 |
| 荧光原位杂交(FISH) | 荧光探针与 rRNA 杂交 | 李斯特菌,沙门氏菌 | 中等 | 数小时 | 细胞的可视化确认 | 样品制备耗时 |
| 微流控检测 | 用于病原体分析的微型通道 | 多种细菌病原体 | 高 | 1 - 3 小时 | 微型化,高通量 | 昂贵,需要微加工 |
| 芯片实验室系统 | 结合多种测定的集成芯片 | 多种病原体同时检测 | 高 | 1 - 2 小时 | 结合检测步骤 | 昂贵,需要熟练操作员 |
| 纳米孔测序 | 单分子测序 | 食源性病原体 | 很高 | 数小时 | 长读长,无扩增 | 错误率高,昂贵 |
| SPR 传感器 | 结合引起的共振信号偏移 | 李斯特菌,沙门氏菌,弧菌 | 高 | 分钟 | 实时监测 | 需要先进设备 |
| 数字 PCR | 分区 DNA 扩增以进行精确定量 | 沙门氏菌(Salmonella)、大肠杆菌(E. coli) | 极高 | 小时级 | 低丰度靶标的精确定量 | 昂贵,需要特殊设备 |
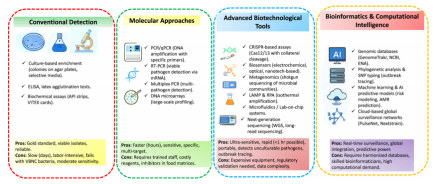
图3.病原体检测技术的演变:从传统和分子方法到先进的生物技术和生物信息学工具,实现了快速、高分辨率的监测
四、防控与干预:绿色与精准策略
生物技术和工程方法正在重新定义食品系统中的病原体控制策略。益生菌和噬菌体提供天然的生物控制,而抗菌肽和工程化细菌素则提供精准靶向的干预措施。表4中提供了关于病原体控制、细菌、抗菌肽、纳米技术、工程微生物和生物聚合物基包装等生物技术和工程策略的综合视图。益生菌、噬菌体、抗菌肽、纳米技术、工程微生物以及食源性病原体控制中生物聚合物包装的机械学基础在图4中被描述。
表4.食品系统中病原体控制的生物技术和工程方法
| 检测方法 | 原理 | 靶标病原体 | 灵敏度 | 检测时间 | 优点 | 缺点 |
|---|---|---|---|---|---|---|
| 基于培养 (Culture-based) | 在选择性/鉴别培养基上生长 | 沙门氏菌, 李斯特菌, 大肠杆菌 | 中等 | 2–7 天 | 金标准,可获得分离株 | 缓慢,漏检VBNC |
| 生化试验 (API, VITEK) | 酶活性,糖发酵 | 肠杆菌科 | 中等 | 1–2 天 | 简单,标准化 | 假阳性/假阴性 |
| PCR/qPCR | 毒力基因的DNA扩增 | 沙门氏菌, 李斯特菌, 大肠杆菌 O157:H7 | 高 (10–100 CFU) | 数小时 | 高灵敏度和特异性 | 易受基质抑制 |
| RT-PCR | RNA转录本检测 | 活性沙门氏菌, 李斯特菌 | 极高 | 数小时 | 检测活病原体 | 昂贵,复杂 |
| 多重 PCR | 多重引物用于多靶标扩增 | 沙门氏菌, 大肠杆菌, 李斯特菌 | 高 | 数小时 | 多病原体检测 | 交叉反应问题 |
| DNA 微阵列 | 探针杂交 | 多种细菌物种 | 高 | 1–2 天 | 大规模分析 | 昂贵,需要先验序列知识 |
| NGS (WGS) | 全基因组测序 | 所有食源性病原体 | 极高 | 1–2 天 | 菌株水平分辨率,疫情追踪 | 成本高,需要生物信息学分析 |
| 宏基因组学 | 所有DNA的鸟枪法测序 | 混合微生物群落 | 高 | 1–3 天 | 检测不可培养病原体 | 昂贵,分析复杂 |
| CRISPR-based 检测 | Cas12/13 旁切活性 | 沙门氏菌, 大肠杆菌, 李斯特菌 | 极高 | <1 小时 | 便携,实时 | 仍为新兴技术 |
| LAMP (环介导等温扩增) | 恒温下的DNA扩增 | 沙门氏菌, 李斯特菌, 弯曲杆菌 | 高 (10 CFU) | 30–60 分钟 | 快速,不需要热循环仪 | 引物设计关键,非特异性扩增 |
| 免疫测定 (ELISA, 侧向流动) | 抗体-抗原结合 | 大肠杆菌, 沙门氏菌, 李斯特菌 | 中等到高 | 1–4 小时 | 廉价,广泛使用 | 可能需要富集,交叉反应 |
| 生物传感器 (电化学, 光学) | 病原体结合后的信号变化 | 沙门氏菌, 李斯特菌, 大肠杆菌, 弧菌 | 高 (单细胞水平可能) | 分钟至2小时 | 便携,灵敏 | 昂贵,需要设备校准 |
| MALDI-TOF 质谱 | 蛋白/肽质量指纹图谱 | 李斯特菌, 沙门氏菌, 大肠杆菌 | 高 | 分钟至数小时 | 快速,准确鉴定 | 依赖数据库,需要分离株 |
| 等温 RPA | 低温下的DNA扩增 | 大肠杆菌, 沙门氏菌 | 高 | 20–40 分钟 | 低温恒温工作 | 对污染敏感 |
| 荧光原位杂交 (FISH) | 荧光探针与rRNA杂交 | 李斯特菌, 沙门氏菌 | 中等 | 数小时 | 细胞的可视化确认 | 样品制备耗时 |
| 微流控检测 | 用于病原体分析的微型通道 | 多种细菌病原体 | 高 | 1–3 小时 | 微型化,高通量 | 昂贵,需要微加工 |
| 芯片实验室系统 | 结合多种测定的集成芯片 | 多种病原体同时检测 | 高 | 1–2 小时 | 结合检测步骤 | 昂贵,需要熟练操作员 |
| 纳米孔测序 | 单分子测序 | 食源性病原体 | 很高 | 数小时 | 长读长,无扩增 | 错误率高,昂贵 |
| SPR 传感器 | 结合引起的共振信号偏移 | 李斯特菌, 沙门氏菌, 弧菌 | 高 | 分钟 | 实时监测 | 需要先进设备 |
| 数字 PCR | 分区 DNA 扩增以进行精确定量 | 沙门氏菌 (Salmonella)、大肠杆菌 (E. coli) | 极高 | 小时级 | 低丰度靶标的精确定量 | 昂贵,需要特殊设备 |
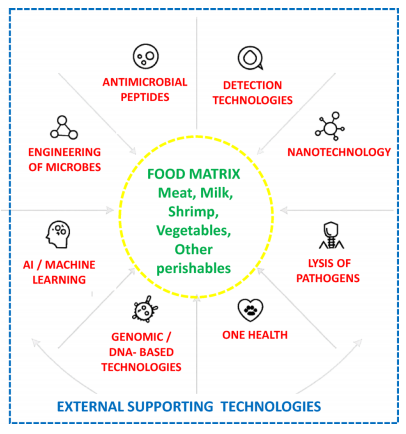
图4.同心示意图,展示了食品基质内部的核心微生物抑制过程,以及能够增强检测、控制和整体食品安全的外部支持技术
4.1 生物防控剂
● 益生菌:乳酸菌 (LAB) 通过竞争排斥、产生细菌素(如乳酸链球菌素)抑制病原菌。例如鼠李糖乳杆菌减少大肠杆菌黏附。
● 噬菌体:高度特异性感染细菌。ListShield™ 和 EcoShield™ 已获批用于控制李斯特菌和大肠杆菌。噬菌体混合制剂可防止细菌产生抗性。
4.2 抗菌肽与纳米技术
● 抗菌肽 (AMPs):破坏细胞膜。工程化改造的细菌素(如乳酸菌素衍生物)对多重耐药菌效果更强。
● 纳米材料:
○ 银纳米颗粒 (AgNPs):破坏细菌呼吸链。
○ 光催化材料:二氧化钛 (TiO₂) 产生活性氧 (ROS) 杀菌。
○ 纳米载体:壳聚糖纳米颗粒包裹精油,实现缓释和增效。
4.3 智能包装与工程菌
● 生物聚合物包装:壳聚糖、海藻酸薄膜结合精油或纳米颗粒,兼具保鲜与抗菌功能。
● 基因工程菌:改造的益生菌(如大肠杆菌 Nissle 1917)可感知病原菌信号并释放抗菌肽,实现"按需"治疗。
五、风险与预防体系
5.1 易感人群与长期后果
● 弱势群体:儿童(免疫未成熟)、老人(免疫力衰退)、孕妇(胎盘感染风险)、免疫缺陷者。
● 后遗症:反应性关节炎、肠易激综合征 (IBS)、肾衰竭等。
5.2 核心预防策略 (Farm-to-Fork)
● 源头控制:良好农业规范 (GAP)。
● 过程管理:危害分析与关键控制点 (HACCP) 体系。
● 冷链管理:保持低温抑制细菌生长(除李斯特菌外)。
● 消费者教育:清洁、生熟分开、彻底煮熟(家禽>74℃)、安全温度储存。
六、挑战与未来展望
6.1 当前面临的瓶颈
● 基质干扰:复杂食品成分(脂肪、蛋白质)抑制检测信号。
● 假阳性/阴性:分子检测可能误报死细胞或漏检低丰度菌。
● 资源鸿沟:WGS和AI技术在低收入国家难以普及。
● 生态风险:纳米材料毒性、噬菌体及工程菌的环境释放安全性需评估。
6.2 未来发展趋势
● 智能化:AI驱动的数字孪生技术模拟污染风险,物联网 (IoT) 实时监控。
● 便携化:低成本、手持式CRISPR/生物传感器进入家庭和农场。
● 多组学整合:基因组、蛋白组、代谢组数据结合,实现对病原体行为的全景预测。
七、结论
食源性病原体的防控正经历从 "被动响应" 向 "主动预测" 的范式转变。通过整合CRISPR诊断、全基因组测序、宏基因组学以及机器学习,我们能够构建更具韧性、数据驱动的食品安全体系。未来的成功关键在于将这些高科技解决方案转化为低成本、可扩展的工具,并加强全球监管协作与数据共享。
参考文献
1.Ahmad S, Lohiya S, Taksande A, Meshram RJ, Varma A, Vagha K, Ahmad S Jr, Sr VA (2024) A comprehensive review of innovative paradigms in microbial detection and antimicrobial resistance: beyond traditional cultural methods. Cureus 16(6):1-8
2.Bhadra L, Dhiman P, Srivastava A, Patel A, Ratrey P, Kumar A, Datta B (2024) Processed food microbiology and safety: Risks, trends and future perspectives. Industrial microbiology and biotechnology: an insight into current trends. Springer Nature Singapore, Singapore, pp 175–227
3.El-Zehery HR, Zaghloul RA, Abdel-Rahman HM, Salem AA, El-Dougdoug KA (2022) Novel strategies of essential oils, chitosan, and nano-chitosan for Inhibition of multi-drug resistant: E. coli O157: H7 and Listeria monocytogenes. Saudi J Biol Sci 29(4):2582–2590
4.Farrukh M, Munawar A, Nawaz Z, Hussain N, Hafeez AB, Szweda P (2025) Antibiotic resistance and preventive strategies in foodborne pathogenic bacteria: A comprehensive review. Food Sci Biotechnol 34(10):2101–2129
5.Ganger S, Harale G, Majumdar P (2023) Clustered regularly interspaced short palindromic repeats/CRISPR-associated (CRISPR/Cas) systems: Discovery, structure, classification, and general mechanism. CRISPR/Cas-Mediated genome editing in plants. Apple Academic, pp 65–97
6.Khoiri S, Moussango VD (2024) A short review on Harnessing bioinformatics for food safety: computational approaches to detecting foodborne pathogens. J Adv Health Inf Res 2(3):109–114
相关产品
HZB221777:肠沙门氏菌肠亚种鸡血清型 | Salmonella enterica subsp. enterica serotype Gallinarum
HZB364569:大肠埃希氏菌 | Escherichia coli O157:H7
HZB355892:单核增生李斯特氏菌 | Listeria monocytogenes (Murray et al.) Pirie
HZB368924:空肠弯曲杆菌亚种 | Campylobacter jejuni subsp. jejuni (Jones et al.) Veron and Chatelains
敬请关注灰藻生物,共筑健康未来!
— 武汉市灰藻生物科技有限公司团队敬上
灰藻生物:我们期待着与客户共同成长,共创生命科学的美好未来!
更新日期:2026-04-10
编制人:思琪
审稿人:小藻